Phospholipid ebb and flow makes mitochondria go
- PMID: 32614384
- PMCID: PMC7401802
- DOI: 10.1083/jcb.202003131
Phospholipid ebb and flow makes mitochondria go
Abstract
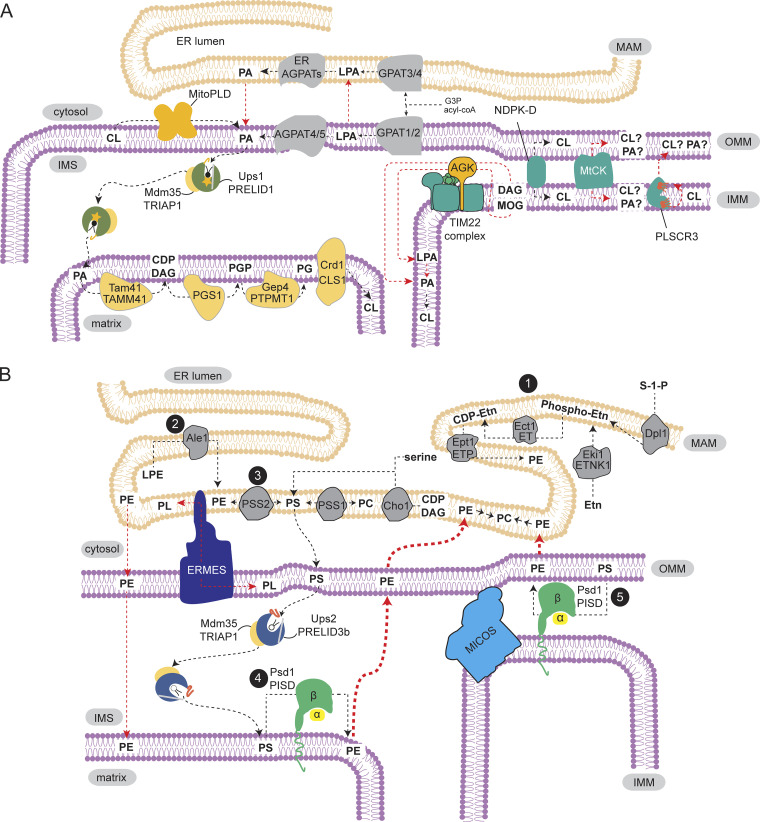
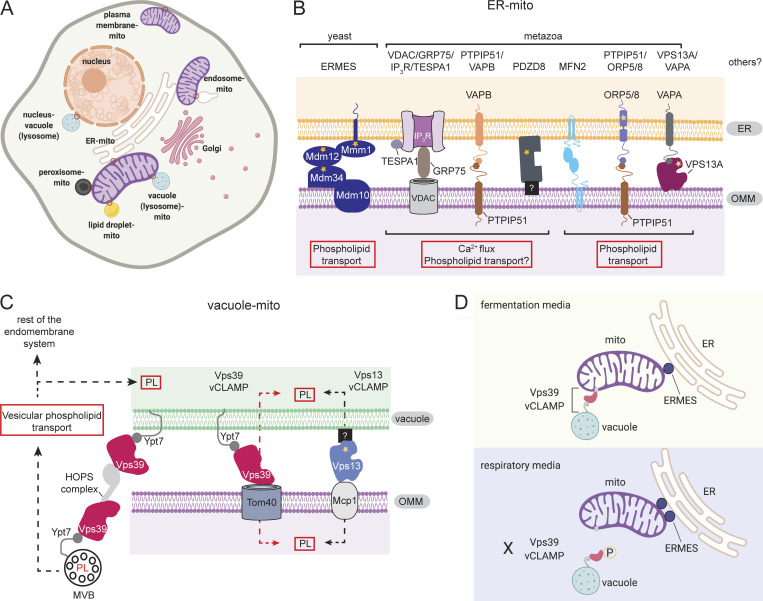
Mitochondria, so much more than just being energy factories, also have the capacity to synthesize macromolecules including phospholipids, particularly cardiolipin (CL) and phosphatidylethanolamine (PE). Phospholipids are vital constituents of mitochondrial membranes, impacting the plethora of functions performed by this organelle. Hence, the orchestrated movement of phospholipids to and from the mitochondrion is essential for cellular integrity. In this review, we capture recent advances in the field of mitochondrial phospholipid biosynthesis and trafficking, highlighting the significance of interorganellar communication, intramitochondrial contact sites, and lipid transfer proteins in maintaining membrane homeostasis. We then discuss the physiological functions of CL and PE, specifically how they associate with protein complexes in mitochondrial membranes to support bioenergetics and maintain mitochondrial architecture.
© 2020 Acoba et al.
Figures