

Single cell heterogeneity in influenza A virus gene expression shapes the innate antiviral response to infection
- PMID: 32614923
- PMCID: PMC7363107
- DOI: 10.1371/journal.ppat.1008671
Single cell heterogeneity in influenza A virus gene expression shapes the innate antiviral response to infection
Abstract
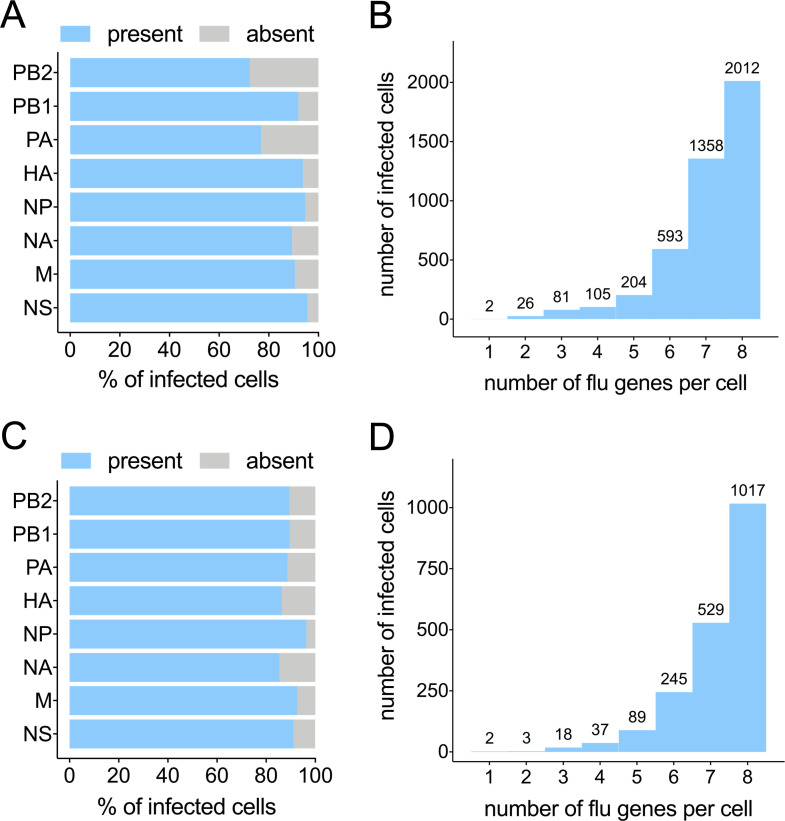
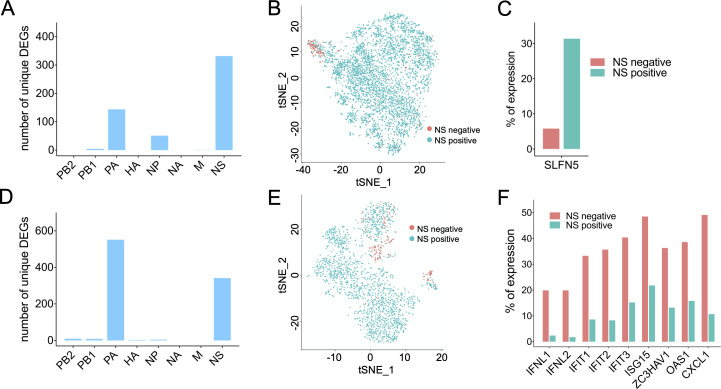
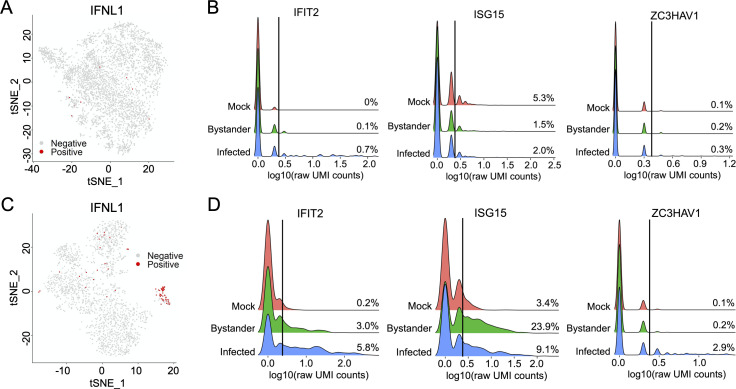
Viral infection outcomes are governed by the complex and dynamic interplay between the infecting virus population and the host response. It is increasingly clear that both viral and host cell populations are highly heterogeneous, but little is known about how this heterogeneity influences infection dynamics or viral pathogenicity. To dissect the interactions between influenza A virus (IAV) and host cell heterogeneity, we examined the combined host and viral transcriptomes of thousands of individual cells, each infected with a single IAV virion. We observed complex patterns of viral gene expression and the existence of multiple distinct host transcriptional responses to infection at the single cell level. We show that human H1N1 and H3N2 strains differ significantly in patterns of both viral and host anti-viral gene transcriptional heterogeneity at the single cell level. Our analyses also reveal that semi-infectious particles that fail to express the viral NS can play a dominant role in triggering the innate anti-viral response to infection. Altogether, these data reveal how patterns of viral population heterogeneity can serve as a major determinant of antiviral gene activation.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
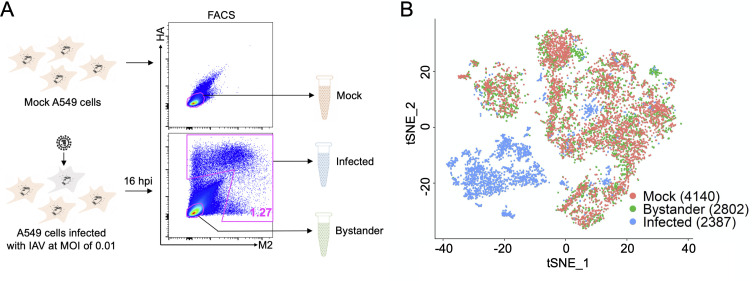
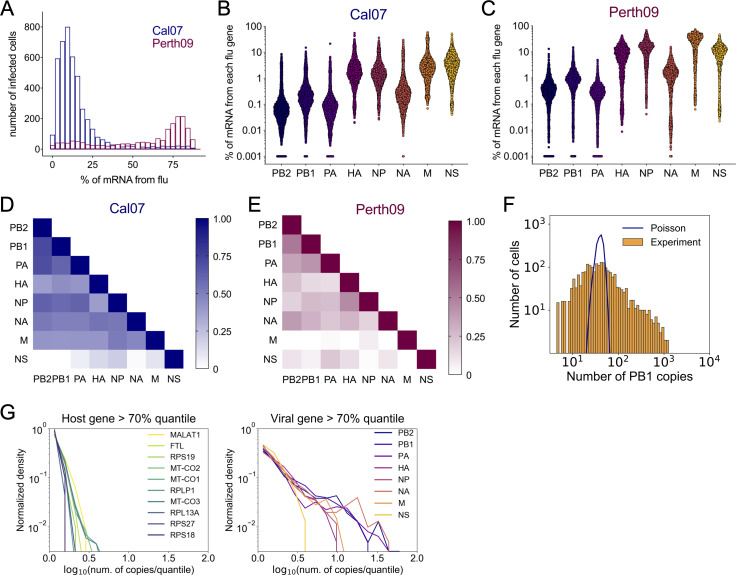
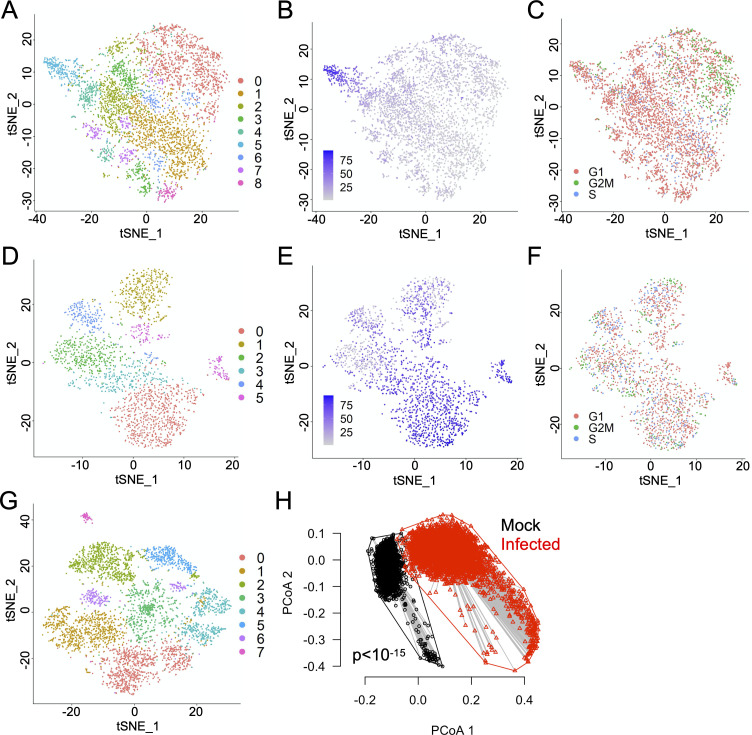
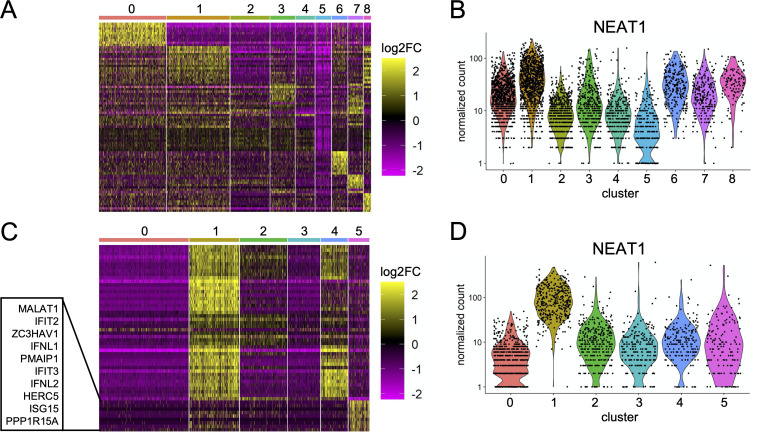
Comment in
-
Influenza virus hedges its bets.Nat Rev Microbiol. 2020 Sep;18(9):475. doi: 10.1038/s41579-020-0422-z. Nat Rev Microbiol. 2020. PMID: 32669680 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
