

Twenty years of t-loops: A case study for the importance of collaboration in molecular biology
- PMID: 32620538
- PMCID: PMC8679138
- DOI: 10.1016/j.dnarep.2020.102901
Twenty years of t-loops: A case study for the importance of collaboration in molecular biology
Abstract
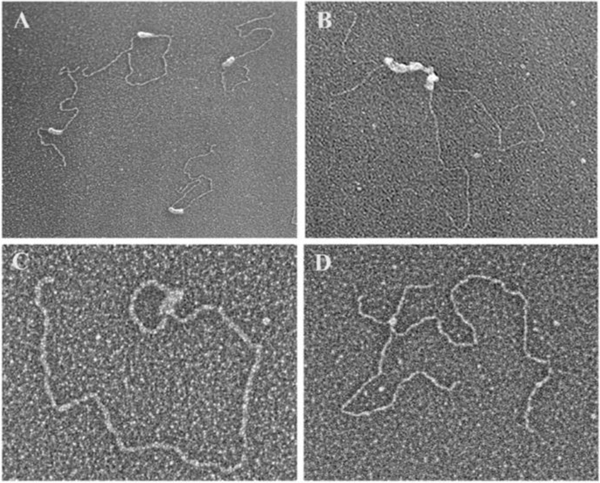
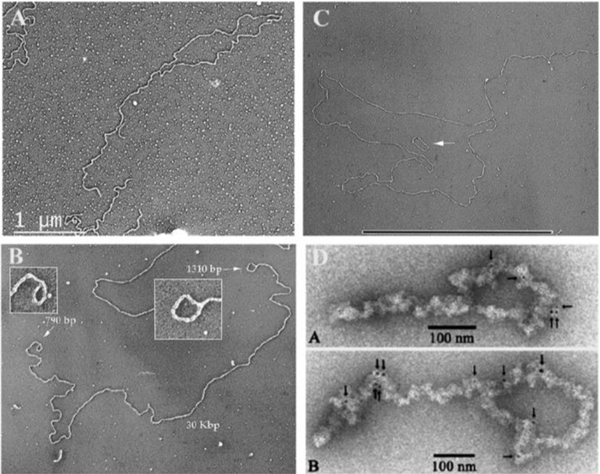
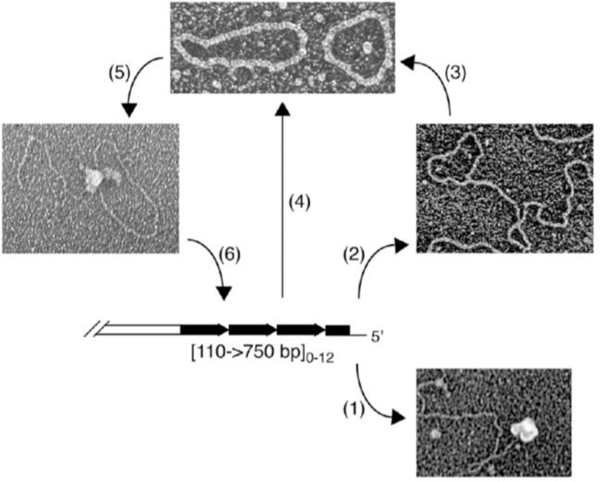
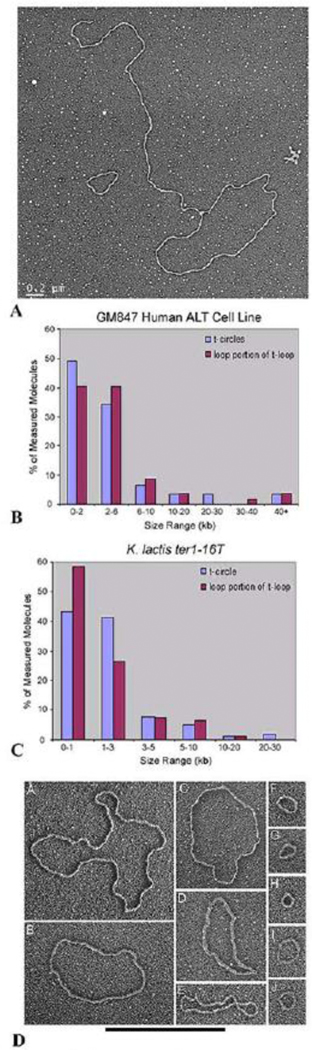
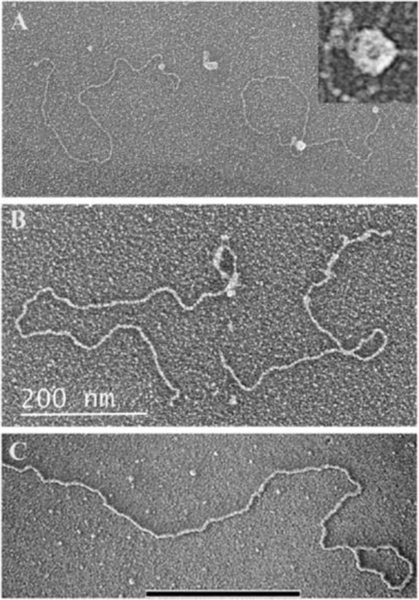
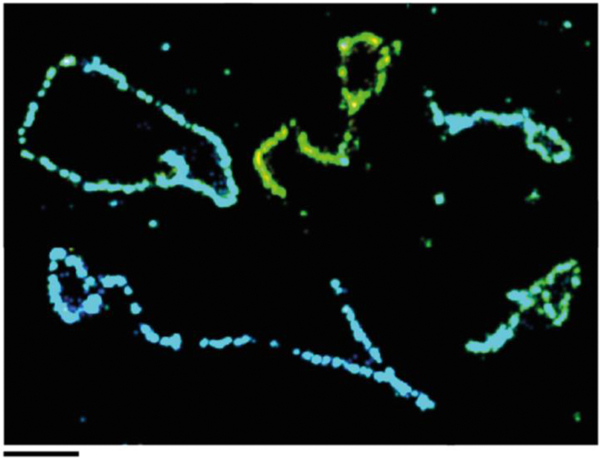
Collaborative studies open doors to breakthroughs otherwise unattainable by any one laboratory alone. Here we describe the initial collaboration between the Griffith and de Lange laboratories that led to thinking about the telomere as a DNA template for homologous recombination, the proposal of telomere looping, and the first electron micrographs of t-loops. This was followed by collaborations that revealed t-loops across eukaryotic phyla. The Griffith and Tomáška/Nosek collaboration revealed circular telomeric DNA (t-circles) derived from the linear mitochondrial chromosomes of nonconventional yeast, which spurred discovery of t-circles in ALT-positive human cells. Collaborative work between the Griffith and McEachern labs demonstrated t-loops and t-circles in a series of yeast species. The de Lange and Zhuang laboratories then applied super-resolution light microscopy to demonstrate a genetic role for TRF2 in loop formation. Recent work from the Griffith laboratory linked telomere transcription with t-loop formation, providing a new model of the t-loop junction. A recent collaboration between the Cesare and Gaus laboratories utilized super-resolution light microscopy to provide details about t-loops as protective elements, followed by the Boulton and Cesare laboratories showing how cell cycle regulation of TRF2 and RTEL enables t-loop opening and reformation to promote telomere replication. Twenty years after the discovery of t-loops, we reflect on the collective history of their research as a case study in collaborative molecular biology.
Keywords: DNA repair; Double strand breaks; R-loop; Super resolution microscopy; T-circle; T-loop; Telomeres.
Copyright © 2020 The Author(s). Published by Elsevier B.V. All rights reserved.
Figures

Comment in
-
Telomeres come to life thanks to an exciting review article in this issue.DNA Repair (Amst). 2020 Oct;94:102904. doi: 10.1016/j.dnarep.2020.102904. Epub 2020 Jun 29. DNA Repair (Amst). 2020. PMID: 32709557 No abstract available.
References
-
- Muller HJ, The remaking of chromosomes. Collecting net, 1938. 13: p. 181–198.
-
- de Lange T, Shelterin-Mediated Telomere Protection. Annu Rev Genet, 2018. 52: p. 223–247. - PubMed
-
- Olovnikov AM, A theory of marginotomy. The incomplete copying of template margin in enzymic synthesis of polynucleotides and biological significance of the phenomenon. J Theor Biol, 1973. 41(1): p. 181–90. - PubMed
-
- Watson JD, Origin of concatemeric T7 DNA. Nat New Biol, 1972. 239(94): p. 197–201. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
