

Bacterial Quorum Sensing Molecules Promote Allergic Airway Inflammation by Activating the Retinoic Acid Response
- PMID: 32622265
- PMCID: PMC7334417
- DOI: 10.1016/j.isci.2020.101288
Bacterial Quorum Sensing Molecules Promote Allergic Airway Inflammation by Activating the Retinoic Acid Response
Abstract
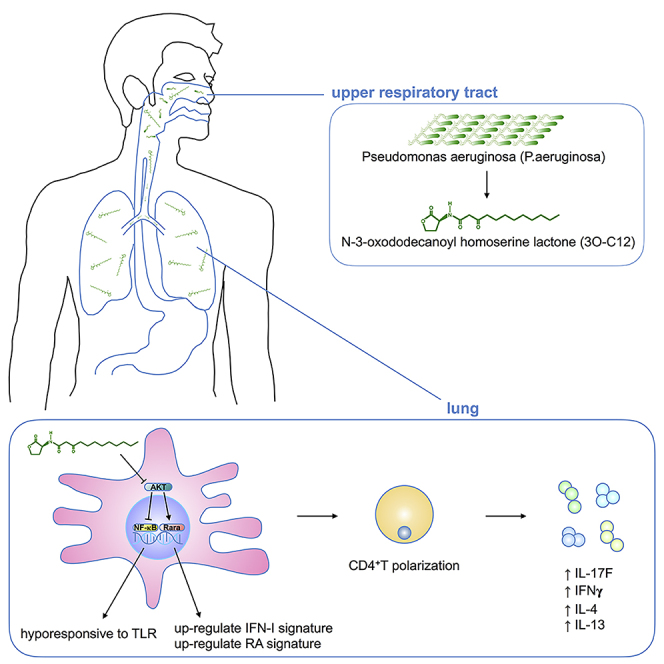
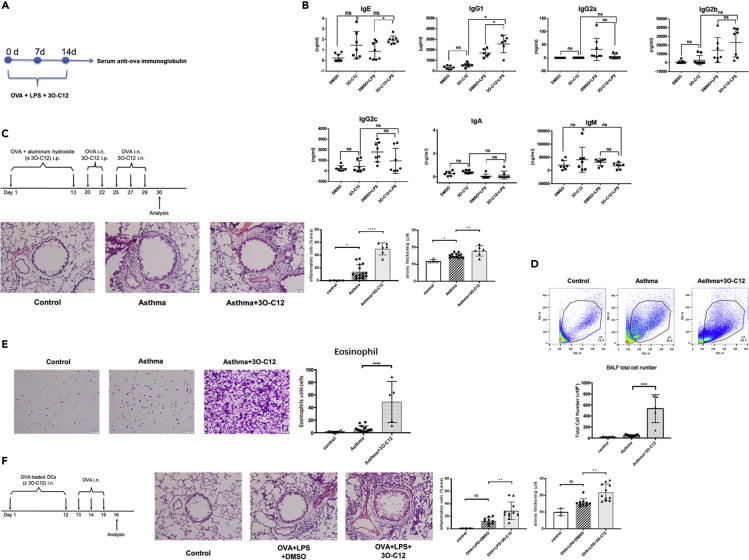
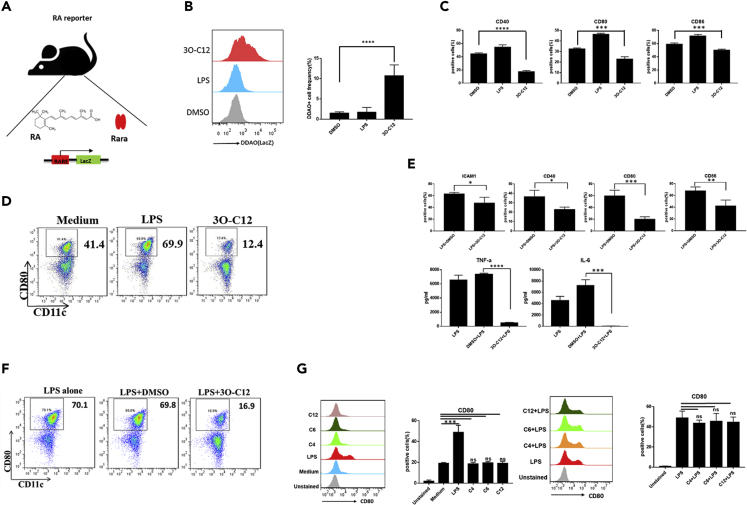
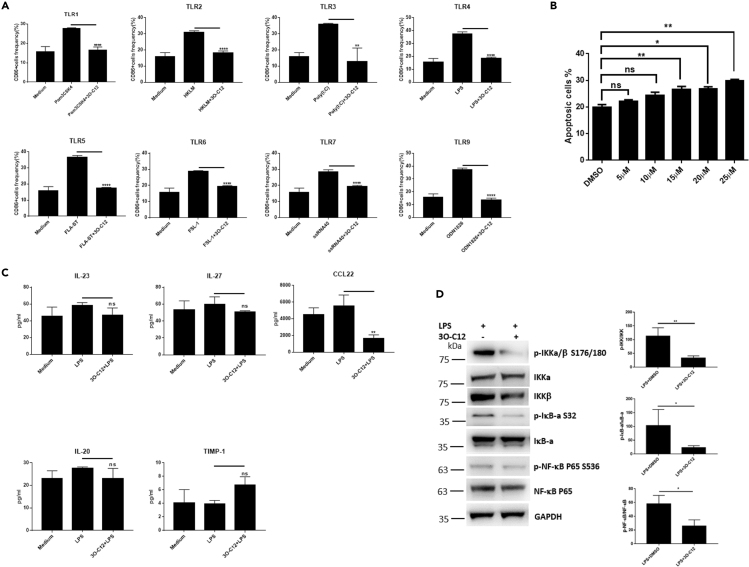
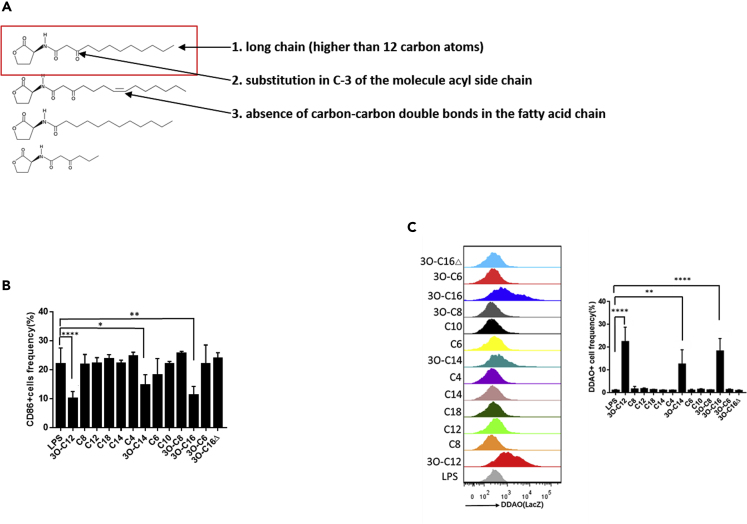
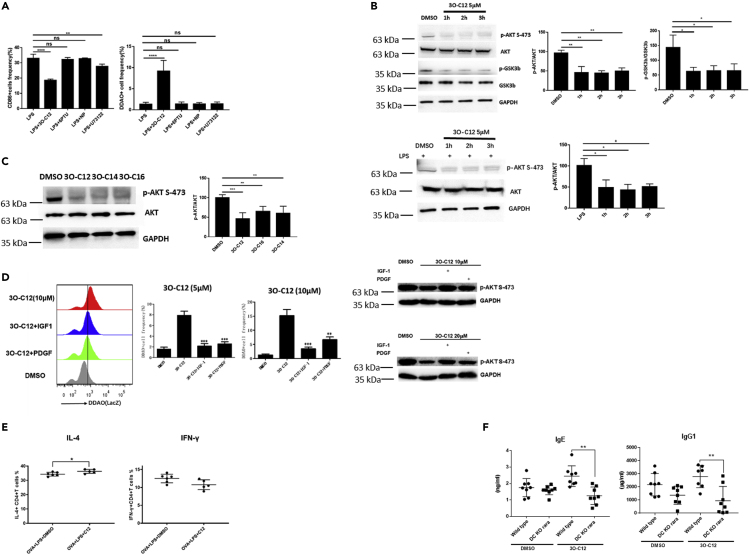
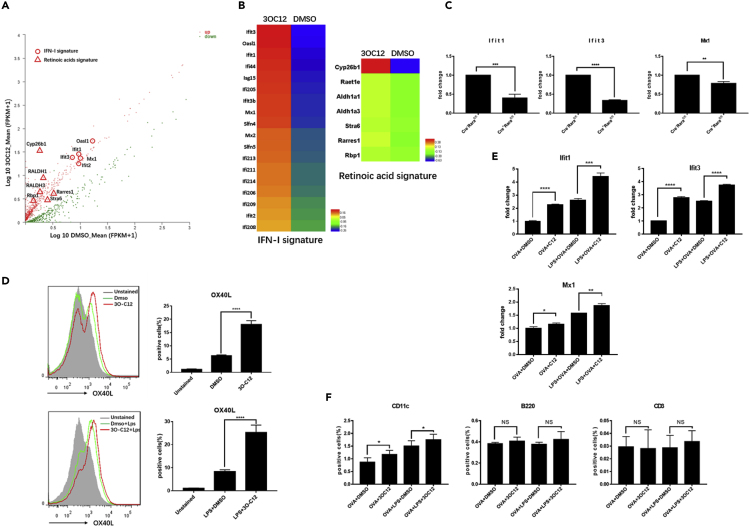
IgE and IgG1 production in the type 2 immune response is the characteristic feature of an allergic reaction. However, whether bacterial molecules modulate IgE and IgG1 production remains obscure. Here, we demonstrate that the bacterial quorum sensing molecules acyl homoserine lactones (AHLs) induce IgE and IgG1 production by activating the RARE (retinoic acid response element) response in dendritic cells (DCs) in vivo. DC-specific knockout of the retinoic acid transcriptional factor Rara diminished the AHL-stimulated type 2 immune response in vitro. AHLs altered DC phenotype, upregulated OX40L and IFN-I signature, and promoted T helper 2 cell differentiation in vitro. Finally, AHLs activated the RARE response by inhibiting AKT phosphorylation in vitro, as the AKT agonists IGF-1 and PDGF abolished the effect of AHLs on the RARE response. This study demonstrates a mechanism by which AHLs drive allergic airway inflammation through activating retinoic acid signaling in DCs.
Keywords: Biological Sciences; Immune Response; Immunology.
Copyright © 2020 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
Similar articles
-
The bile acid-activated retinoic acid response in dendritic cells is involved in food allergen sensitization.Allergy. 2022 Feb;77(2):483-498. doi: 10.1111/all.15039. Epub 2021 Aug 14. Allergy. 2022. PMID: 34365653
-
Bacillus megaterium CYP102A1 oxidation of acyl homoserine lactones and acyl homoserines.Biochemistry. 2007 Dec 18;46(50):14429-37. doi: 10.1021/bi701945j. Epub 2007 Nov 20. Biochemistry. 2007. PMID: 18020460
-
Identification of Quorum-Sensing Molecules of N-Acyl-Homoserine Lactone in Gluconacetobacter Strains by Liquid Chromatography-Tandem Mass Spectrometry.Molecules. 2019 Jul 24;24(15):2694. doi: 10.3390/molecules24152694. Molecules. 2019. PMID: 31344938 Free PMC article.
-
Beneficial effects of bacteria-plant communication based on quorum sensing molecules of the N-acyl homoserine lactone group.Plant Mol Biol. 2016 Apr;90(6):605-12. doi: 10.1007/s11103-016-0457-8. Epub 2016 Feb 22. Plant Mol Biol. 2016. PMID: 26898296 Review.
-
Detection of bacterial quorum sensing N-acyl homoserine lactones in clinical samples.Anal Bioanal Chem. 2008 Jul;391(5):1619-27. doi: 10.1007/s00216-008-2002-3. Epub 2008 Apr 12. Anal Bioanal Chem. 2008. PMID: 18408921 Review.
Cited by
-
Gossip in the gut: Quorum sensing, a new player in the host-microbiota interactions.World J Gastroenterol. 2021 Nov 14;27(42):7247-7270. doi: 10.3748/wjg.v27.i42.7247. World J Gastroenterol. 2021. PMID: 34876787 Free PMC article. Review.
-
Short-Chain Fatty Acids Calibrate RARα Activity Regulating Food Sensitization.Front Immunol. 2021 Oct 14;12:737658. doi: 10.3389/fimmu.2021.737658. eCollection 2021. Front Immunol. 2021. PMID: 34721398 Free PMC article.
-
Biological Function of Antimicrobial Peptides on Suppressing Pathogens and Improving Host Immunity.Antibiotics (Basel). 2023 Jun 10;12(6):1037. doi: 10.3390/antibiotics12061037. Antibiotics (Basel). 2023. PMID: 37370356 Free PMC article. Review.
-
Antimicrobial Peptides and Their Biomedical Applications: A Review.Antibiotics (Basel). 2024 Aug 23;13(9):794. doi: 10.3390/antibiotics13090794. Antibiotics (Basel). 2024. PMID: 39334969 Free PMC article. Review.
References
-
- Bachus H., Kaur K., Papillion A.M., Marquez-Lago T.T., Yu Z., Ballesteros-Tato A., Matalon S., Leon B. Impaired tumor-necrosis-factor-alpha-driven dendritic cell activation limits lipopolysaccharide-induced protection from allergic inflammation in infants. Immunity. 2019;50:225–240.e4. - PMC - PubMed
-
- Bhattacharya N., Yuan R., Prestwood T.R., Penny H.L., DiMaio M.A., Reticker-Flynn N.E., Krois C.R., Kenkel J.A., Pham T.D., Carmi Y. Normalizing microbiota-induced retinoic acid deficiency stimulates protective CD8(+) T cell-mediated immunity in colorectal cancer. Immunity. 2016;45:641–655. - PMC - PubMed
-
- Bisgaard H., Li N., Bonnelykke K., Chawes B.L., Skov T., Paludan-Muller G., Stokholm J., Smith B., Krogfelt K.A. Reduced diversity of the intestinal microbiota during infancy is associated with increased risk of allergic disease at school age. J. Allergy Clin. Immunol. 2011;128:646–652.e1-5. - PubMed
-
- Cait A., Hughes M.R., Antignano F., Cait J., Dimitriu P.A., Maas K.R., Reynolds L.A., Hacker L., Mohr J., Finlay B.B. Microbiome-driven allergic lung inflammation is ameliorated by short-chain fatty acids. Mucosal Immunol. 2018;11:785–795. - PubMed
-
- Chen C., Perry T.L., Chitko-McKown C.G., Smith A.D., Cheung L., Beshah E., Urban J.F., Jr., Dawson H.D. The regulatory actions of retinoic acid on M2 polarization of porcine macrophages. Dev. Comp. Immunol. 2019;98:20–33. - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
