

Tying up loose ends: the N-degron and C-degron pathways of protein degradation
- PMID: 32627813
- PMCID: PMC7458402
- DOI: 10.1042/BST20191094
Tying up loose ends: the N-degron and C-degron pathways of protein degradation
Abstract
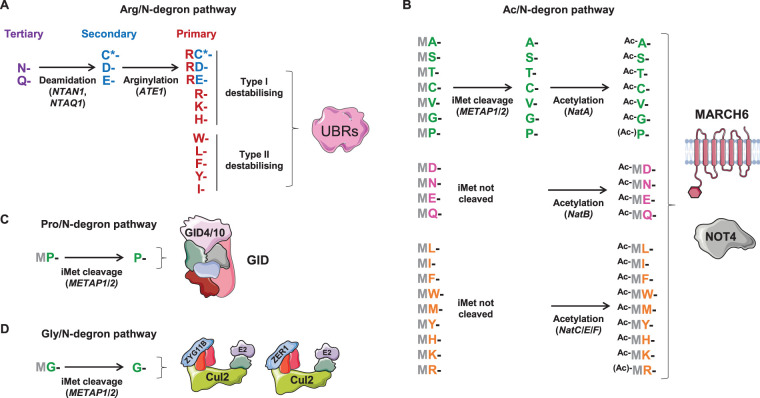
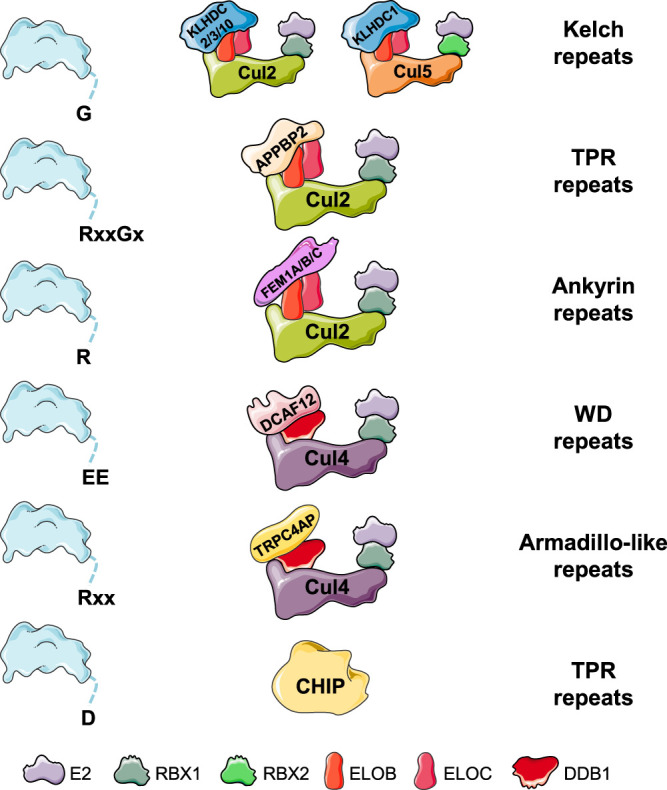
Selective protein degradation by the ubiquitin-proteasome system (UPS) is thought to be governed primarily by the recognition of specific motifs - degrons - present in substrate proteins. The ends of proteins - the N- and C-termini - have unique properties, and an important subset of protein-protein interactions involve the recognition of free termini. The first degrons to be discovered were located at the extreme N-terminus of proteins, a finding which initiated the study of the N-degron (formerly N-end rule) pathways, but only in the last few years has it emerged that a diverse set of C-degron pathways target analogous degron motifs located at the extreme C-terminus of proteins. In this minireview we summarise the N-degron and C-degron pathways currently known to operate in human cells, focussing primarily on those that have been discovered in recent years. In each case we describe the cellular machinery responsible for terminal degron recognition, and then consider some of the functional roles of terminal degron pathways. Altogether, a broad spectrum of E3 ubiquitin ligases mediate the recognition of a diverse array of terminal degron motifs; these degradative pathways have the potential to influence a wide variety of cellular functions.
Keywords: C-degron pathways; E3 ubiquitin ligases; N-degron pathways; degron; protein termini; ubiquitin proteasome system.
© 2020 The Author(s).
Conflict of interest statement
The authors declare that there are no competing interests associated with the manuscript.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources