

CTCF as a boundary factor for cohesin-mediated loop extrusion: evidence for a multi-step mechanism
- PMID: 32631111
- PMCID: PMC7566886
- DOI: 10.1080/19491034.2020.1782024
CTCF as a boundary factor for cohesin-mediated loop extrusion: evidence for a multi-step mechanism
Abstract
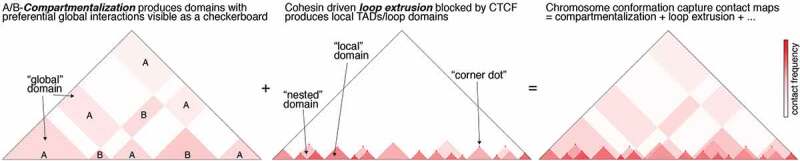
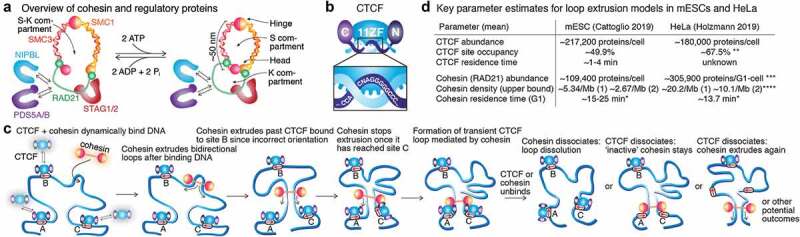
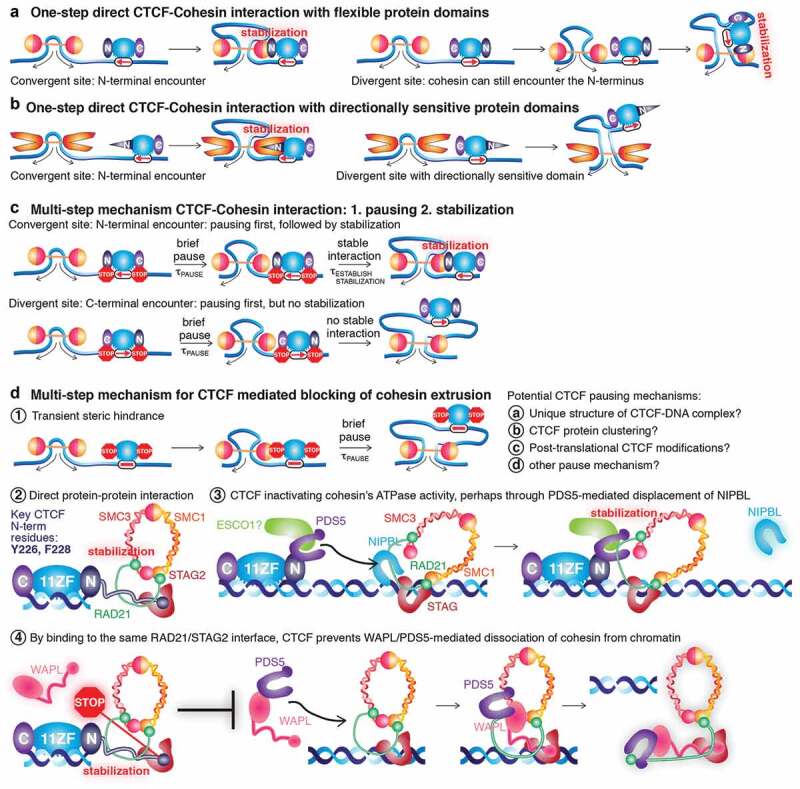
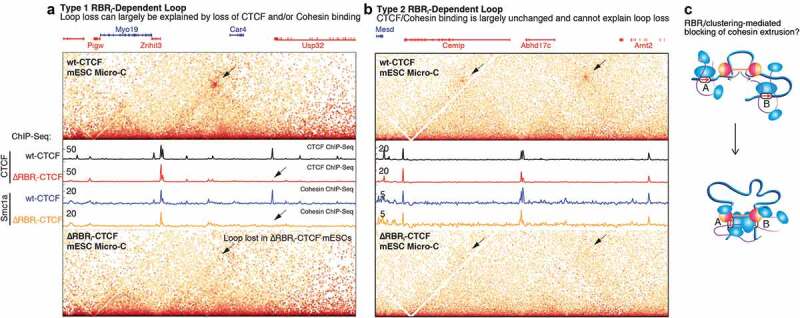
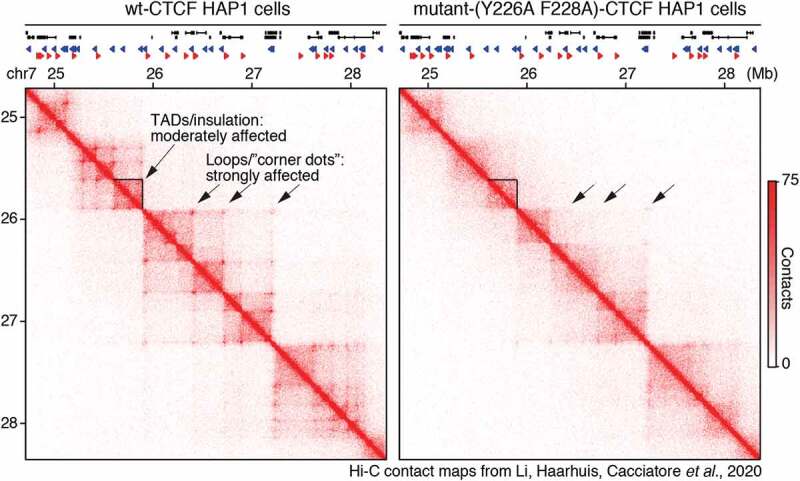
Mammalian genome structure is closely linked to function. At the scale of kilobases to megabases, CTCF and cohesin organize the genome into chromatin loops. Mechanistically, cohesin is proposed to extrude chromatin loops bidirectionally until it encounters occupied CTCF DNA-binding sites. Curiously, loops form predominantly between CTCF binding sites in a convergent orientation. How CTCF interacts with and blocks cohesin extrusion in an orientation-specific manner has remained a mechanistic mystery. Here, we review recent papers that have shed light on these processes and suggest a multi-step interaction between CTCF and cohesin. This interaction may first involve a pausing step, where CTCF halts cohesin extrusion, followed by a stabilization step of the CTCF-cohesin complex, resulting in a chromatin loop. Finally, we discuss our own recent studies on an internal RNA-Binding Region (RBRi) in CTCF to elucidate its role in regulating CTCF clustering, target search mechanisms and chromatin loop formation and future challenges.
Keywords: Binding Polarity; CTCF; Cohesin; Convergent Rule; Loop Extrusion; NIPBL; PDS5; RNA-Binding Region; TADs.
Conflict of interest statement
I declare that no competing financial interests exist.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials