

Origin, conservation, and loss of alternative splicing events that diversify the proteome in Saccharomycotina budding yeasts
- PMID: 32631843
- PMCID: PMC7491326
- DOI: 10.1261/rna.075655.120
Origin, conservation, and loss of alternative splicing events that diversify the proteome in Saccharomycotina budding yeasts
Abstract
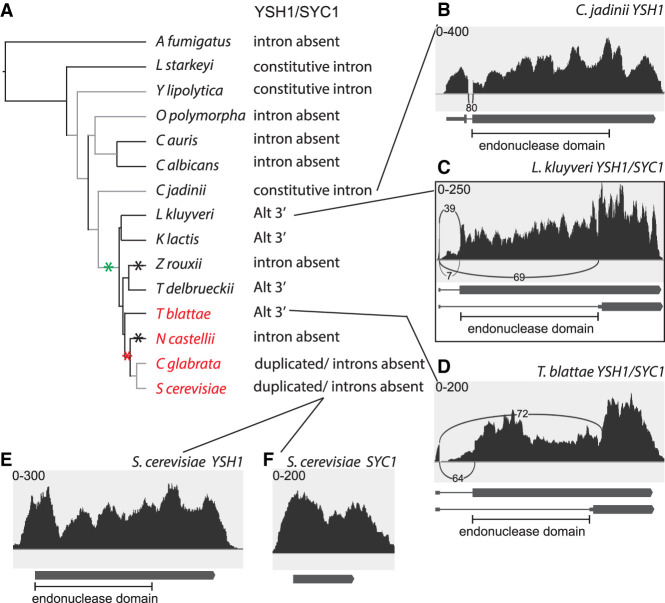
Many eukaryotes use RNA processing, including alternative splicing, to express multiple gene products from the same gene. The budding yeast Saccharomyces cerevisiae has been successfully used to study the mechanism of splicing and the splicing machinery, but alternative splicing in yeast is relatively rare and has not been extensively studied. Alternative splicing of SKI7/HBS1 is widely conserved, but yeast and a few other eukaryotes have replaced this one alternatively spliced gene with a pair of duplicated, unspliced genes as part of a whole genome doubling (WGD). We show that other examples of alternative splicing known to have functional consequences are widely conserved within Saccharomycotina. A common mechanism by which alternative splicing has disappeared is by replacement of an alternatively spliced gene with duplicate unspliced genes. This loss of alternative splicing does not always take place soon after duplication, but can take place after sufficient time has elapsed for speciation. Saccharomycetaceae that diverged before WGD use alternative splicing more frequently than S. cerevisiae, suggesting that WGD is a major reason for infrequent alternative splicing in yeast. We anticipate that WGDs in other lineages may have had the same effect. Having observed that two functionally distinct splice-isoforms are often replaced by duplicated genes allowed us to reverse the reasoning. We thereby identify several splice isoforms that are likely to produce two functionally distinct proteins because we find them replaced by duplicated genes in related species. We also identify some alternative splicing events that are not conserved in closely related species and unlikely to produce functionally distinct proteins.
Keywords: alternative splicing; evolution; subfunctionalization; yeast.
© 2020 Hurtig et al.; Published by Cold Spring Harbor Laboratory Press for the RNA Society.
Figures










References
-
- Awad AM, Venkataramanan S, Nag A, Galivanche AR, Bradley MC, Neves LT, Douglass S, Clarke CF, Johnson TL. 2017. Chromatin-remodeling SWI/SNF complex regulates coenzyme Q6 synthesis and a metabolic shift to respiration in yeast. J Biol Chem 292: 14851–14866. 10.1074/jbc.M117.798397 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases