

The emergent interactions that govern biodiversity change
- PMID: 32632009
- PMCID: PMC7382255
- DOI: 10.1073/pnas.2003852117
The emergent interactions that govern biodiversity change
Abstract
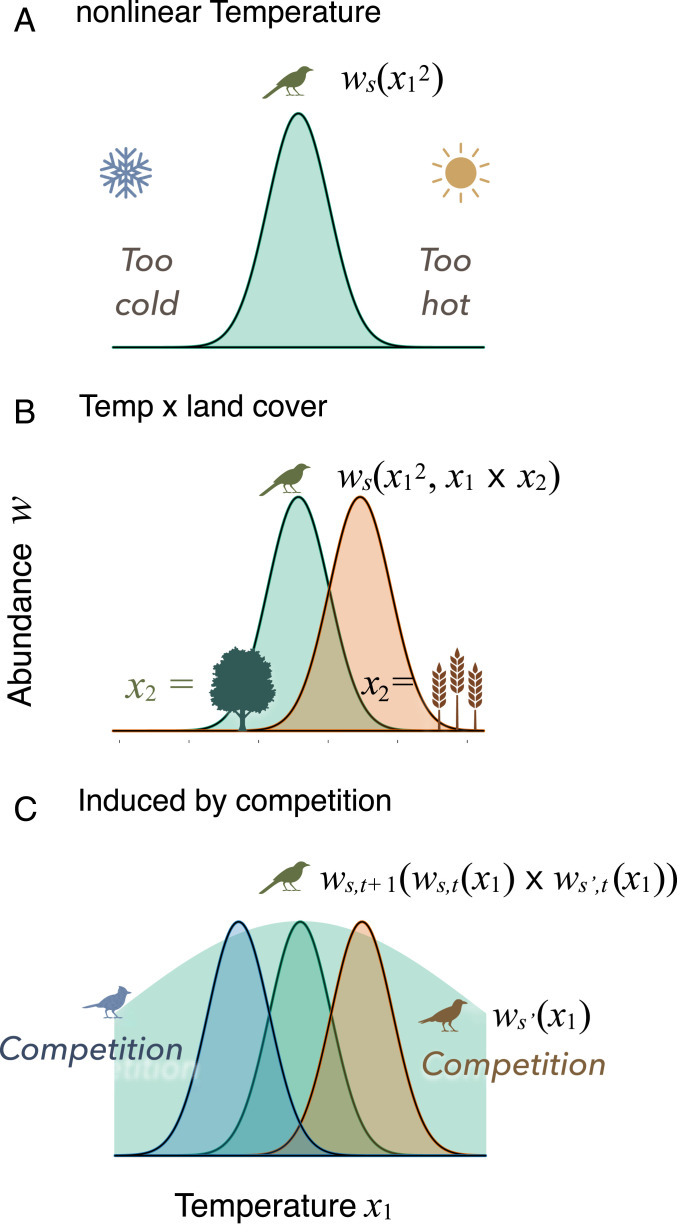
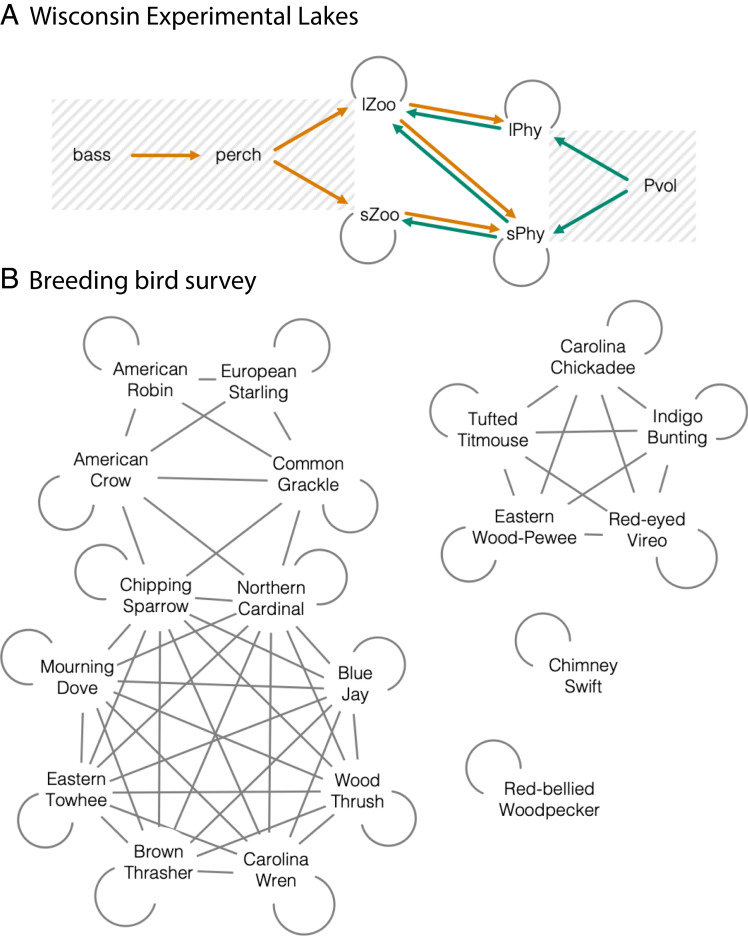
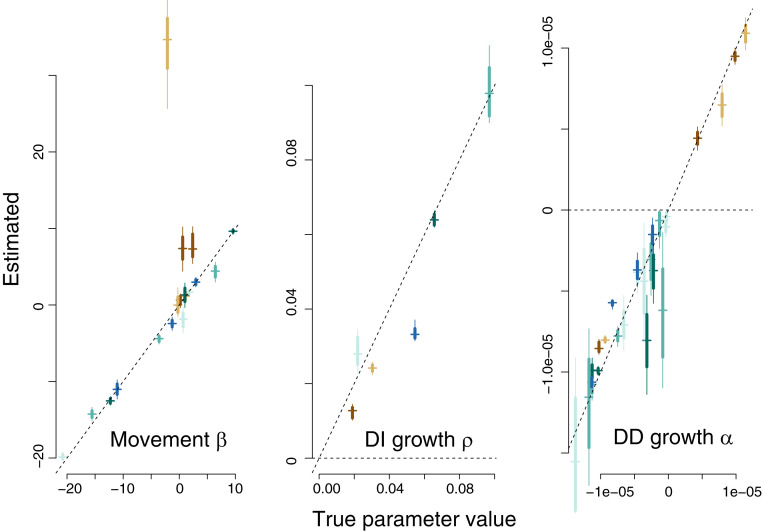
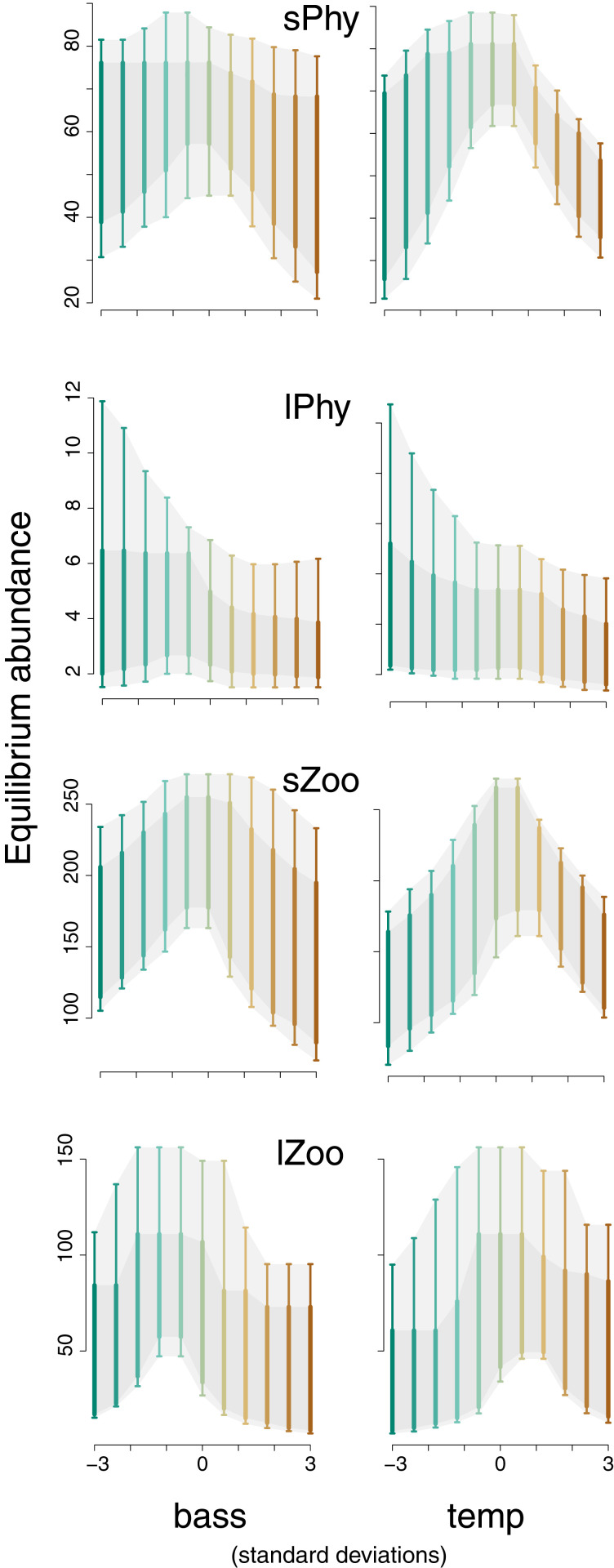
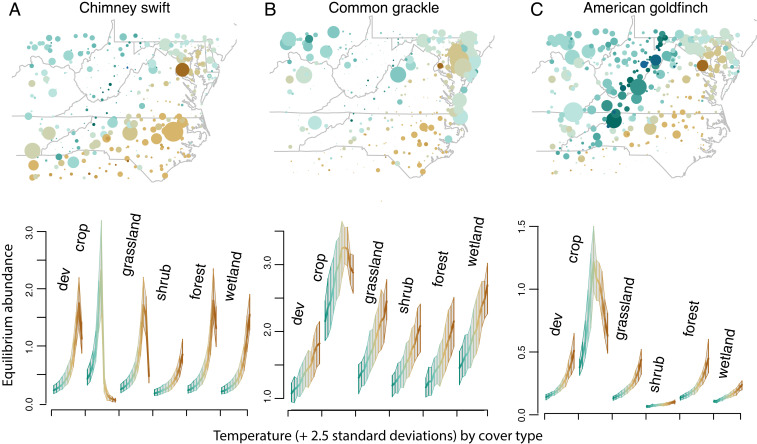
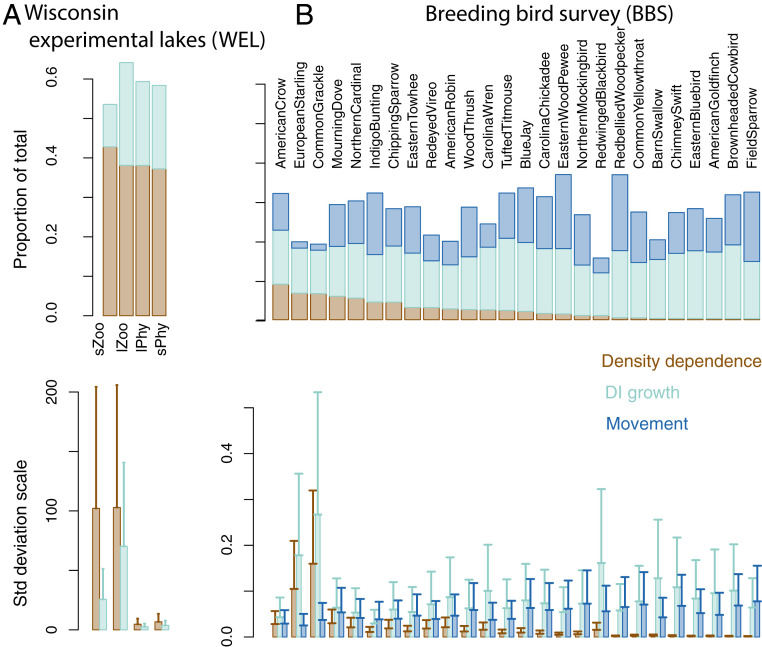
Observational studies have not yet shown that environmental variables can explain pervasive nonlinear patterns of species abundance, because those patterns could result from (indirect) interactions with other species (e.g., competition), and models only estimate direct responses. The experiments that could extract these indirect effects at regional to continental scales are not feasible. Here, a biophysical approach quantifies environment- species interactions (ESI) that govern community change from field data. Just as species interactions depend on population abundances, so too do the effects of environment, as when drought is amplified by competition. By embedding dynamic ESI within framework that admits data gathered on different scales, we quantify responses that are induced indirectly through other species, including probabilistic uncertainty in parameters, model specification, and data. Simulation demonstrates that ESI are needed for accurate interpretation. Analysis demonstrates how nonlinear responses arise even when their direct responses to environment are linear. Applications to experimental lakes and the Breeding Bird Survey (BBS) yield contrasting estimates of ESI. In closed lakes, interactions involving phytoplankton and their zooplankton grazers play a large role. By contrast, ESI are weak in BBS, as expected where year-to-year movement degrades the link between local population growth and species interactions. In both cases, nonlinear responses to environmental gradients are induced by interactions between species. Stability analysis indicates stability in the closed-system lakes and instability in BBS. The probabilistic framework has direct application to conservation planning that must weigh risk assessments for entire habitats and communities against competing interests.
Keywords: GJAM; climate change; food web dynamics; species interactions.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Hutchinson G., Concluding remarks. Cold Spring Harbor Symp. Quant. Biol. 22, 415–427 (1957).
-
- Paine R. T., Food webs—Road maps of interactions or grist for theoretical development. Ecology 69, 1648–1654 (1988).
-
- Novak M., et al. , Predicting community responses to perturbations in the face of imperfect knowledge and network complexity. Ecology 92, 836–846 (2011). - PubMed
-
- Calabrese J. M., Certain G., Kraan C., Dormann C. F., Stacking species distribution models and adjusting bias by linking them to macroecological models. Global Ecol. Biogeogr. 23, 99–112 (2014).
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
