

How frequency hopping suppresses pulse-echo ambiguity in bat biosonar
- PMID: 32632013
- PMCID: PMC7382275
- DOI: 10.1073/pnas.2001105117
How frequency hopping suppresses pulse-echo ambiguity in bat biosonar
Abstract
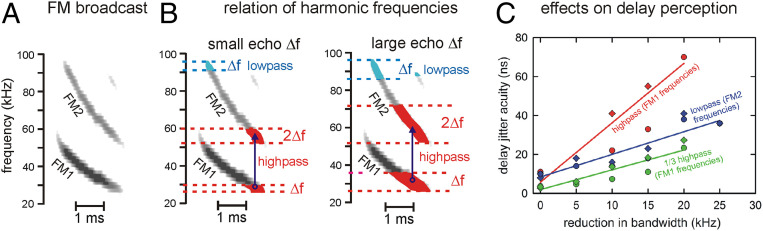
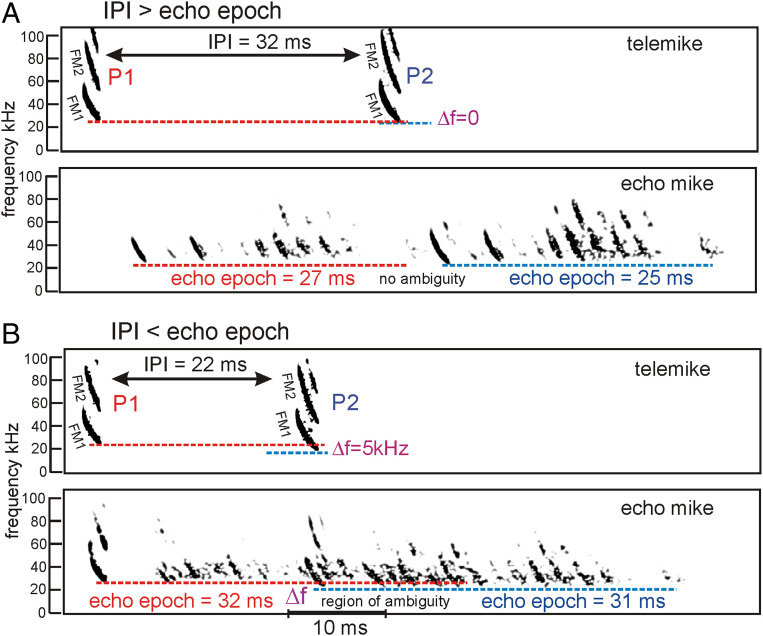
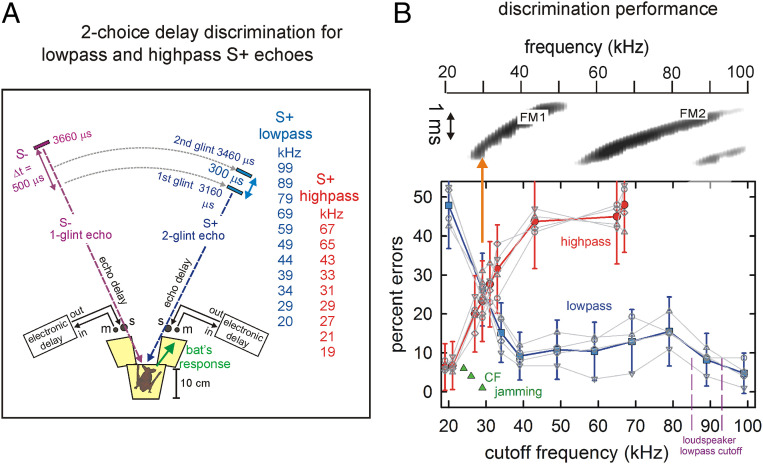
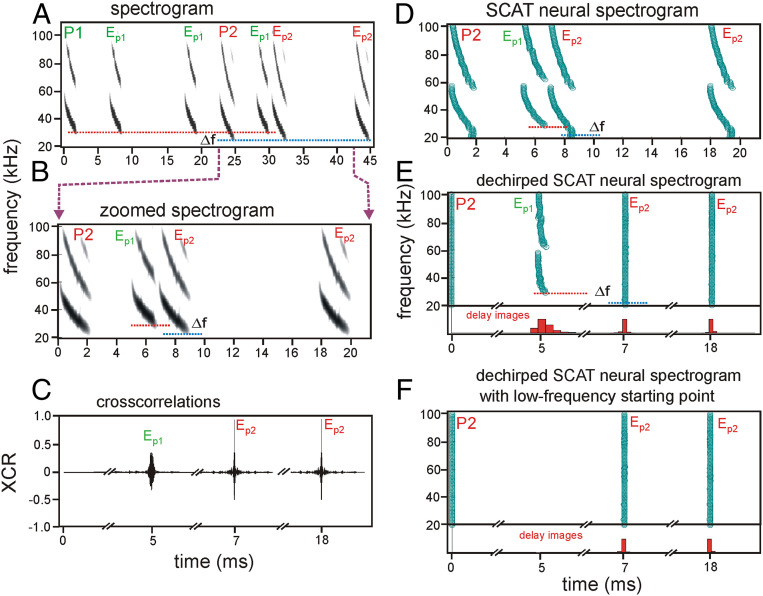
Big brown bats transmit wideband FM biosonar sounds that sweep from 55 to 25 kHz (first harmonic, FM1) and from 110 to 50 kHz (second harmonic, FM2). FM1 is required to perceive echo delay for target ranging; FM2 contributes only if corresponding FM1 frequencies are present. We show that echoes need only the lowest FM1 broadcast frequencies of 25 to 30 kHz for delay perception. If these frequencies are removed, no delay is perceived. Bats begin echo processing at the lowest frequencies and accumulate perceptual acuity over successively higher frequencies, but they cannot proceed without the low-frequency starting point in their broadcasts. This reveals a solution to pulse-echo ambiguity, a serious problem for radar or sonar. In dense, extended biosonar scenes, bats have to emit sounds rapidly to avoid collisions with near objects. But if a new broadcast is emitted when echoes of the previous broadcast still are arriving, echoes from both broadcasts intermingle, creating ambiguity about which echo corresponds to which broadcast. Frequency hopping by several kilohertz from one broadcast to the next can segregate overlapping narrowband echo streams, but wideband FM echoes ordinarily do not segregate because their spectra still overlap. By starting echo processing at the lowest frequencies in frequency-hopped broadcasts, echoes of the higher hopped broadcast are prevented from being accepted by lower hopped broadcasts, and ambiguity is avoided. The bat-inspired spectrogram correlation and transformation (SCAT) model also begins at the lowest frequencies; echoes that lack them are eliminated from processing of delay and no longer cause ambiguity.
Keywords: bat biosonar; clutter suppression; echo ambiguity; echolocation; sonar image.
Copyright © 2020 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
Similar articles
-
FM echolocating bats shift frequencies to avoid broadcast-echo ambiguity in clutter.Proc Natl Acad Sci U S A. 2010 Apr 13;107(15):7048-53. doi: 10.1073/pnas.1000429107. Epub 2010 Mar 29. Proc Natl Acad Sci U S A. 2010. PMID: 20351291 Free PMC article.
-
Biosonar interpulse intervals and pulse-echo ambiguity in four species of echolocating bats.J Exp Biol. 2019 Apr 15;222(Pt 8):jeb195446. doi: 10.1242/jeb.195446. J Exp Biol. 2019. PMID: 30877230
-
Perception of echo delay is disrupted by small temporal misalignment of echo harmonics in bat sonar.J Exp Biol. 2011 Feb 1;214(Pt 3):394-401. doi: 10.1242/jeb.048983. J Exp Biol. 2011. PMID: 21228198 Free PMC article.
-
Temporal binding of neural responses for focused attention in biosonar.J Exp Biol. 2014 Aug 15;217(Pt 16):2834-43. doi: 10.1242/jeb.104380. J Exp Biol. 2014. PMID: 25122915 Free PMC article. Review.
-
Bats use a neuronally implemented computational acoustic model to form sonar images.Curr Opin Neurobiol. 2012 Apr;22(2):311-9. doi: 10.1016/j.conb.2012.02.007. Epub 2012 Mar 19. Curr Opin Neurobiol. 2012. PMID: 22436892 Review.
Cited by
-
A comprehensive computational model of animal biosonar signal processing.PLoS Comput Biol. 2021 Feb 17;17(2):e1008677. doi: 10.1371/journal.pcbi.1008677. eCollection 2021 Feb. PLoS Comput Biol. 2021. PMID: 33596199 Free PMC article.
-
Analysis of echolocation behavior of bats in "echo space" using acoustic simulation.BMC Biol. 2022 Mar 14;20(1):59. doi: 10.1186/s12915-022-01253-y. BMC Biol. 2022. PMID: 35282831 Free PMC article.
-
Echolocating Bats Have Evolved Decreased Susceptibility to Noise-Induced Temporary Hearing Losses.J Assoc Res Otolaryngol. 2024 Jun;25(3):229-238. doi: 10.1007/s10162-024-00941-6. Epub 2024 Apr 2. J Assoc Res Otolaryngol. 2024. PMID: 38565735 Free PMC article. Review.
References
-
- Kurta A., Baker R. H., Eptesicus fuscus. Mamm. Species 356, 1–10 (1990).
-
- Griffin D. R., Listening in the Dark, (Yale University Press, New Haven, CT, 1958).
-
- Neuweiler G., Biology of Bats, (Oxford University Press, Oxford, UK, 2000).
-
- Fenton M. B., Grinnell A. D., Popper A. N., Eds., Bat Bioacoustics, (Springer, New York, 2016).
-
- Clare E. L., Symondson W. O. C., Fenton M. B., An inordinate fondness for beatles? Variation in seasonal dietary preferences of night-roosting big brown bats (Eptesicus fuscus). Mol. Ecol. 23, 3633–3647 (2014). - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
