

RNA contributions to the form and function of biomolecular condensates
- PMID: 32632317
- PMCID: PMC7785677
- DOI: 10.1038/s41580-020-0264-6
RNA contributions to the form and function of biomolecular condensates
Abstract
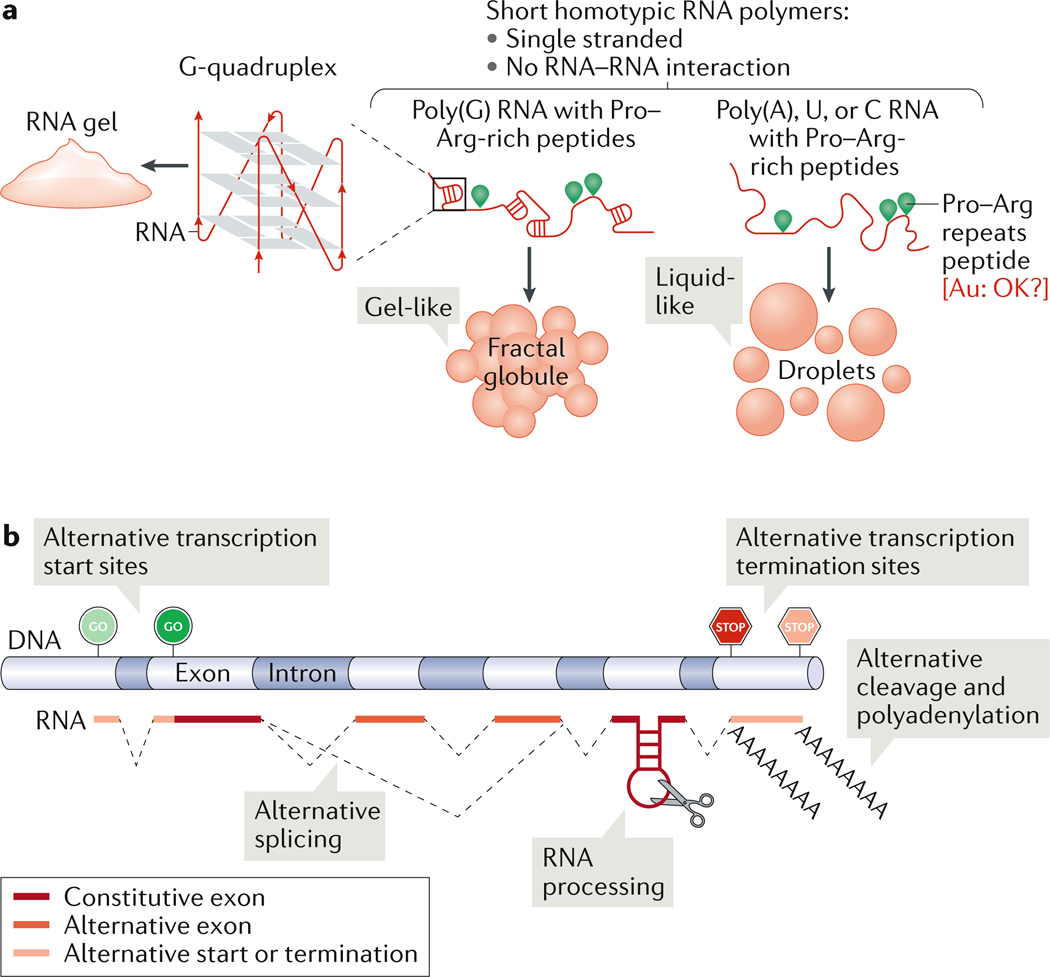
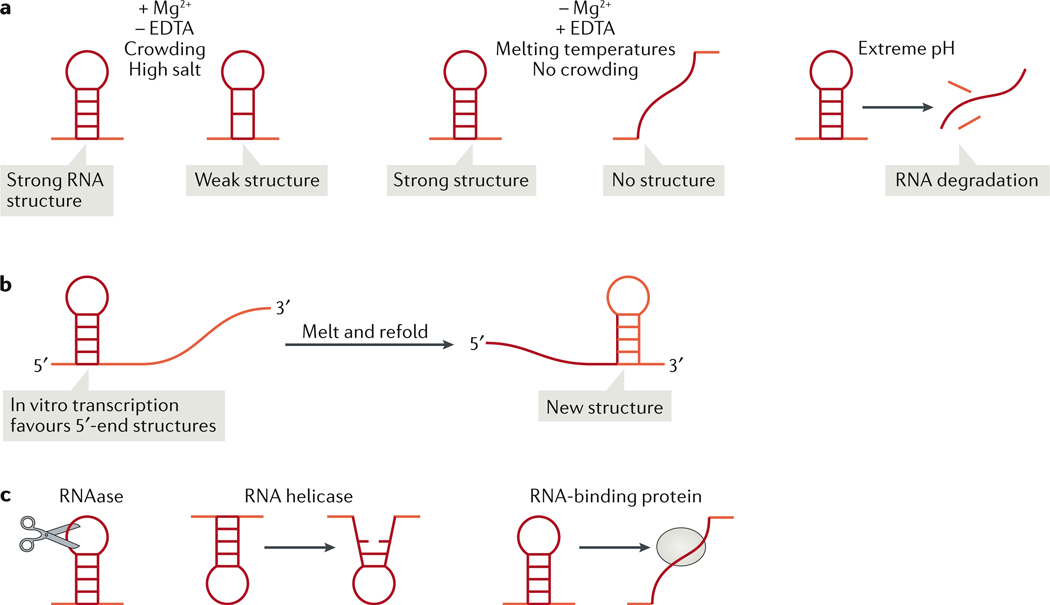
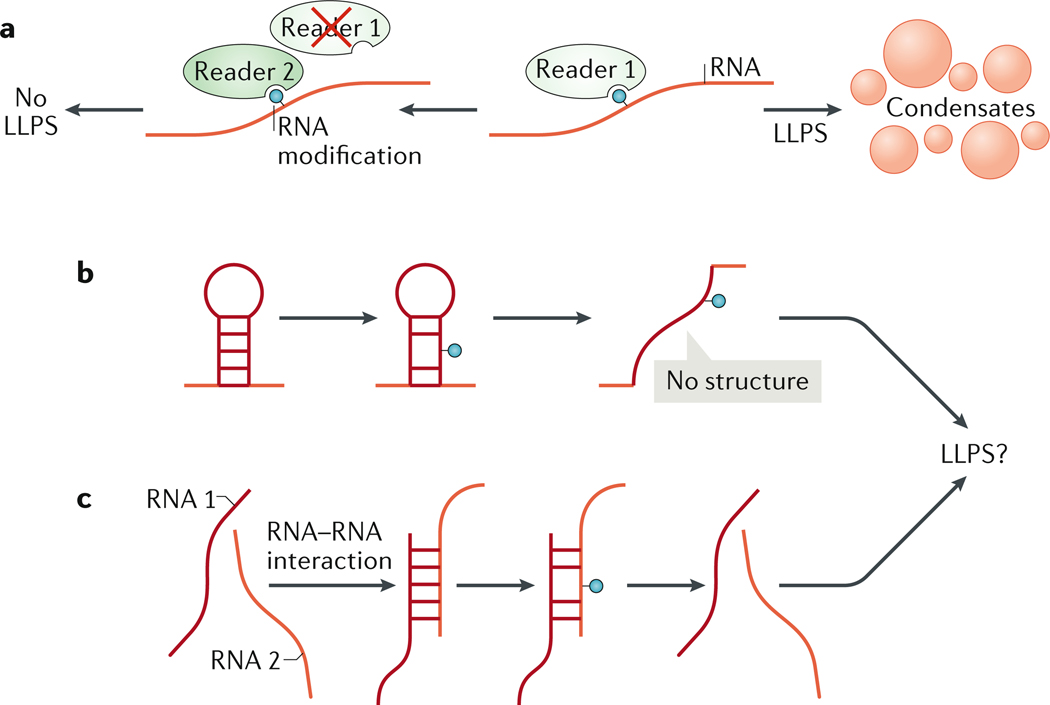
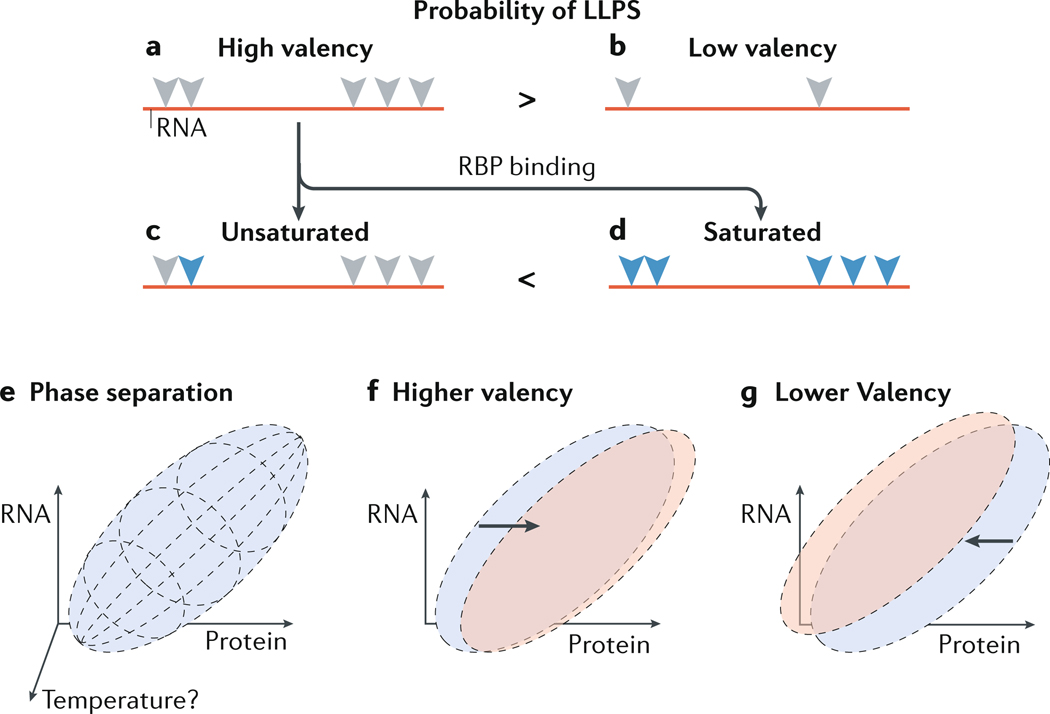
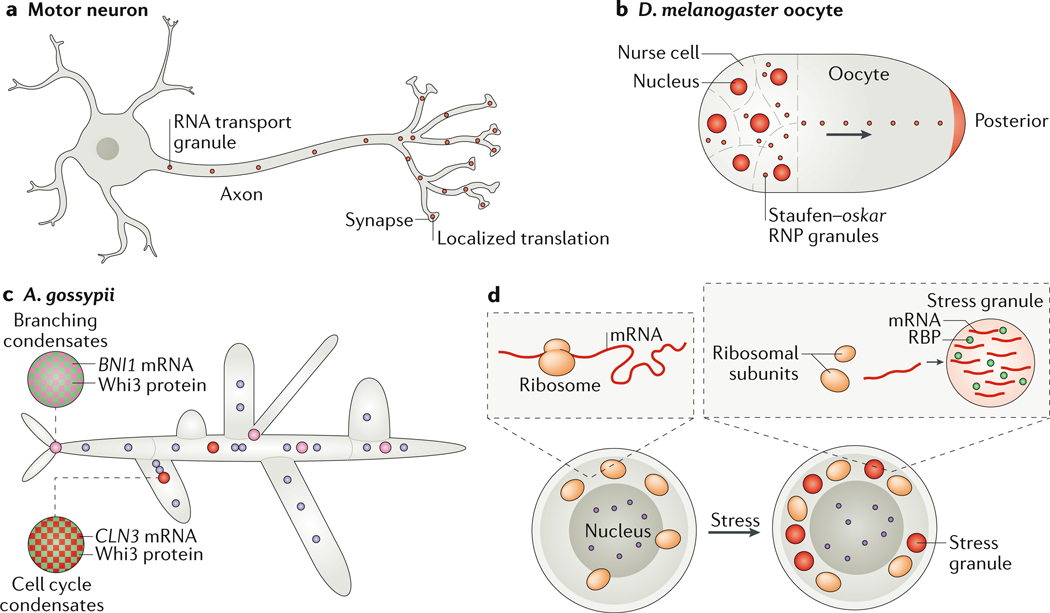
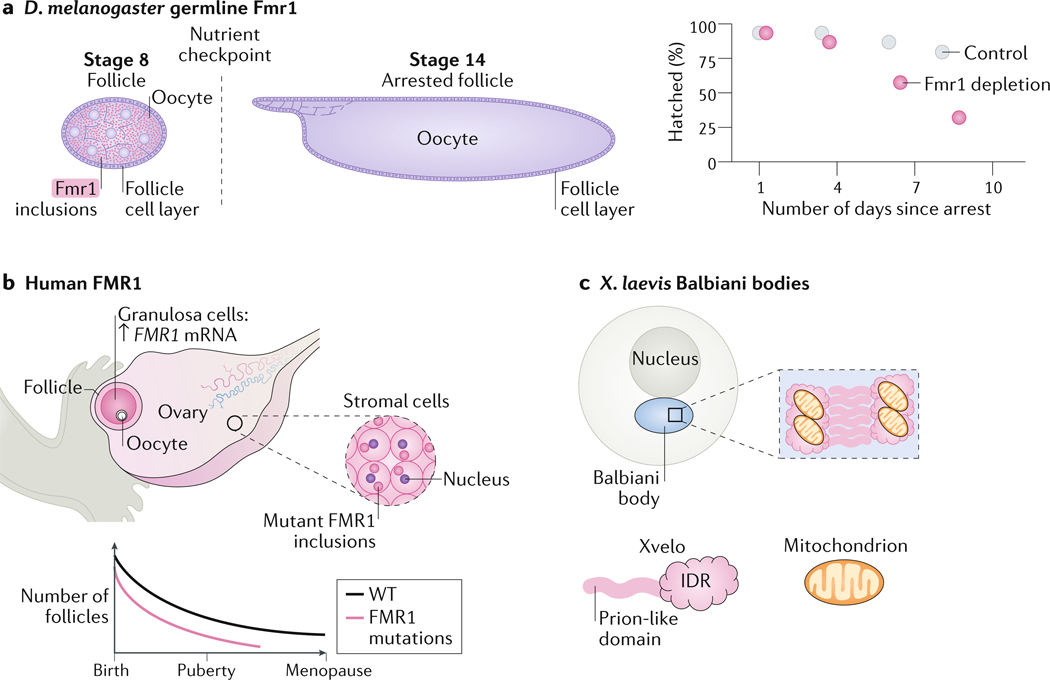
Biomolecular condensation partitions cellular contents and has important roles in stress responses, maintaining homeostasis, development and disease. Many nuclear and cytoplasmic condensates are rich in RNA and RNA-binding proteins (RBPs), which undergo liquid-liquid phase separation (LLPS). Whereas the role of RBPs in condensates has been well studied, less attention has been paid to the contribution of RNA to LLPS. In this Review, we discuss the role of RNA in biomolecular condensation and highlight considerations for designing condensate reconstitution experiments. We focus on RNA properties such as composition, length, structure, modifications and expression level. These properties can modulate the biophysical features of native condensates, including their size, shape, viscosity, liquidity, surface tension and composition. We also discuss the role of RNA-protein condensates in development, disease and homeostasis, emphasizing how their properties and function can be determined by RNA. Finally, we discuss the multifaceted cellular functions of biomolecular condensates, including cell compartmentalization through RNA transport and localization, supporting catalytic processes, storage and inheritance of specific molecules, and buffering noise and responding to stress.
Conflict of interest statement
Conflict of Interest:
The authors declare no competing financial interests
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
