

A plant DNA virus replicates in the salivary glands of its insect vector via recruitment of host DNA synthesis machinery
- PMID: 32636269
- PMCID: PMC7382290
- DOI: 10.1073/pnas.1820132117
A plant DNA virus replicates in the salivary glands of its insect vector via recruitment of host DNA synthesis machinery
Abstract
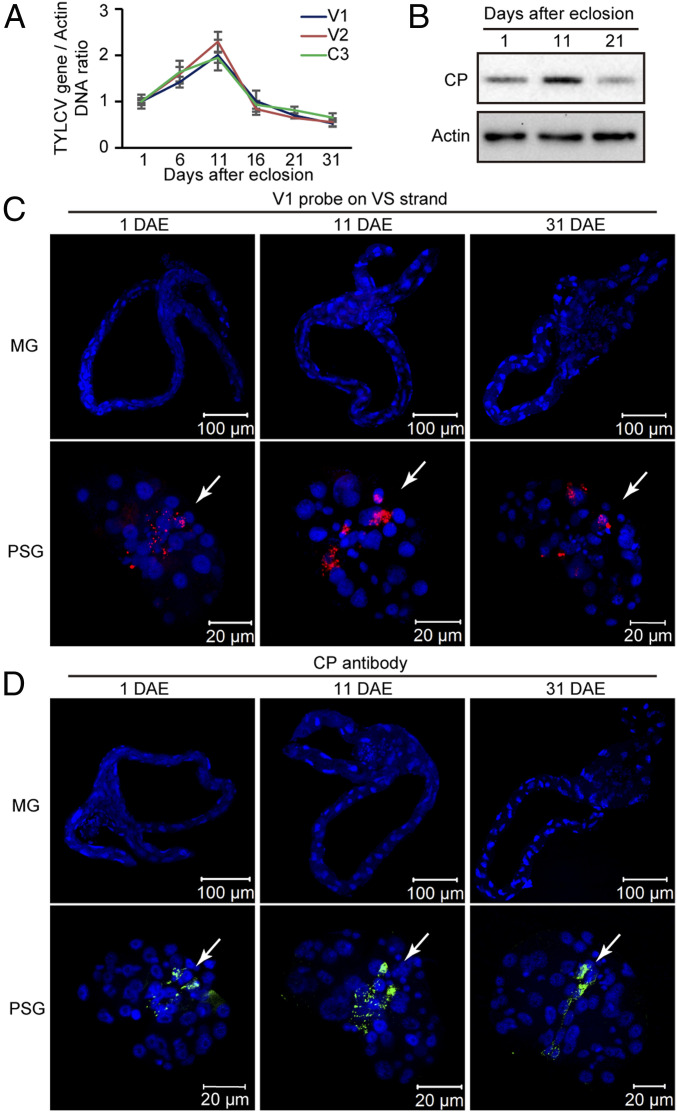
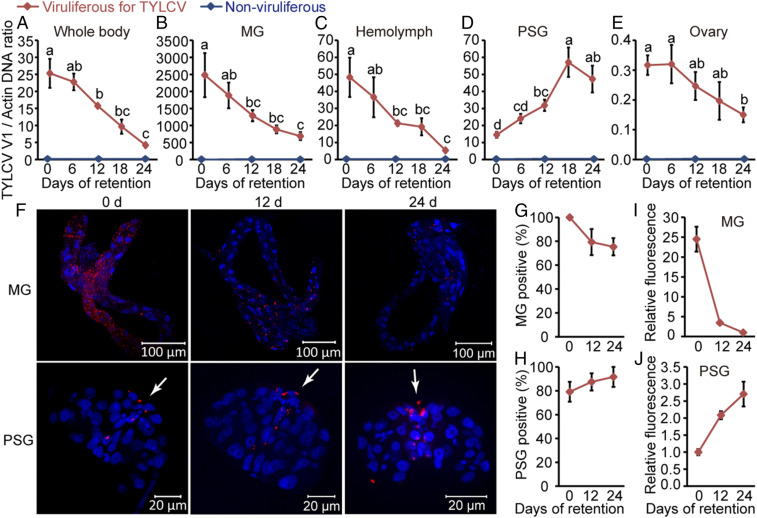
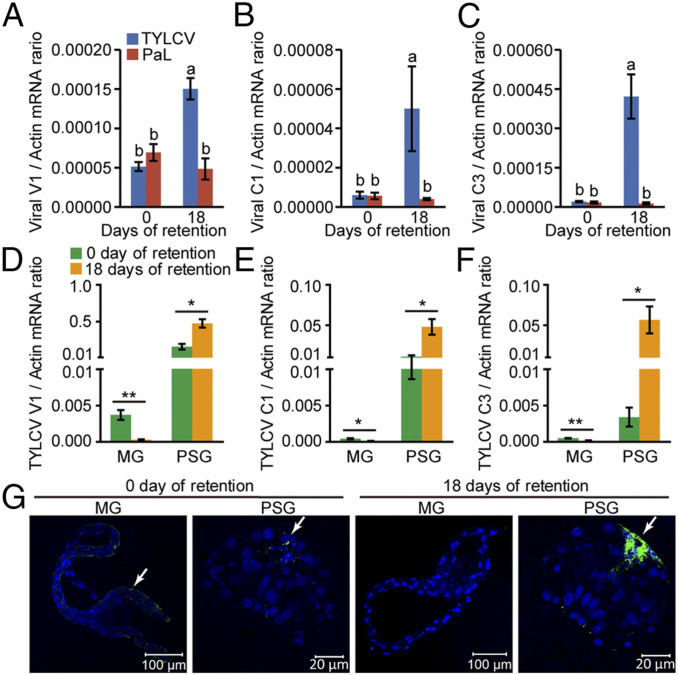
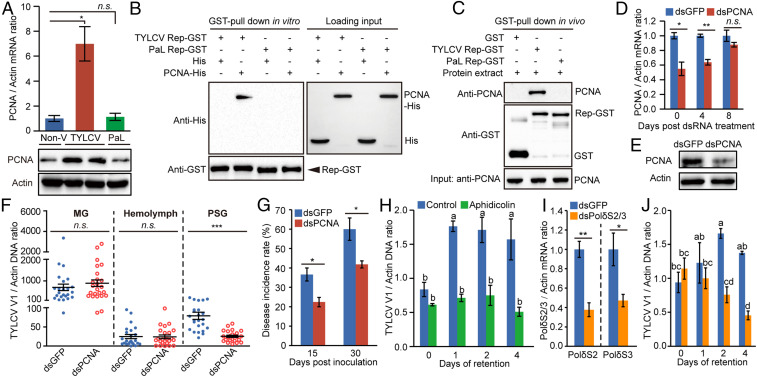
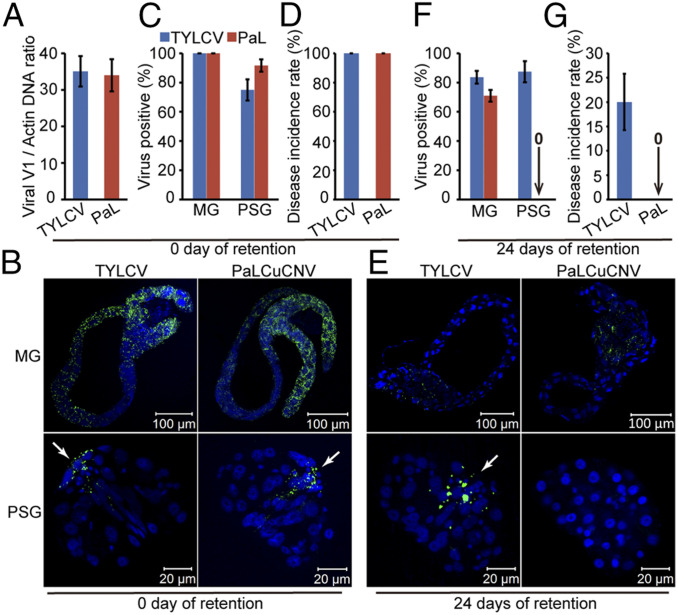
Whereas most of the arthropod-borne animal viruses replicate in their vectors, this is less common for plant viruses. So far, only some plant RNA viruses have been demonstrated to replicate in insect vectors and plant hosts. How plant viruses evolved to replicate in the animal kingdom remains largely unknown. Geminiviruses comprise a large family of plant-infecting, single-stranded DNA viruses that cause serious crop losses worldwide. Here, we report evidence and insight into the replication of the geminivirus tomato yellow leaf curl virus (TYLCV) in the whitefly (Bemisia tabaci) vector and that replication is mainly in the salivary glands. We found that TYLCV induces DNA synthesis machinery, proliferating cell nuclear antigen (PCNA) and DNA polymerase δ (Polδ), to establish a replication-competent environment in whiteflies. TYLCV replication-associated protein (Rep) interacts with whitefly PCNA, which recruits DNA Polδ for virus replication. In contrast, another geminivirus, papaya leaf curl China virus (PaLCuCNV), does not replicate in the whitefly vector. PaLCuCNV does not induce DNA-synthesis machinery, and the Rep does not interact with whitefly PCNA. Our findings reveal important mechanisms by which a plant DNA virus replicates across the kingdom barrier in an insect and may help to explain the global spread of this devastating pathogen.
Keywords: DNA synthesis machinery; insect vector; plant DNA virus; replication; salivary glands.
Copyright © 2020 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Eigenbrode S. D., Bosque-Pérez N. A., Davis T. S., Insect-borne plant pathogens and their vectors: Ecology, evolution, and complex interactions. Annu. Rev. Entomol. 63, 169–191 (2018). - PubMed
-
- Hogenhout S. A., Ammar D., Whitfield A. E., Redinbaugh M. G., Insect vector interactions with persistently transmitted viruses. Annu. Rev. Phytopathol. 46, 327–359 (2008). - PubMed
-
- Ng J. C. K., Falk B. W., Virus-vector interactions mediating nonpersistent and semipersistent transmission of plant viruses. Annu. Rev. Phytopathol. 44, 183–212 (2006). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
