

A redox-active switch in fructosamine-3-kinases expands the regulatory repertoire of the protein kinase superfamily
- PMID: 32636308
- PMCID: PMC8455029
- DOI: 10.1126/scisignal.aax6313
A redox-active switch in fructosamine-3-kinases expands the regulatory repertoire of the protein kinase superfamily
Abstract
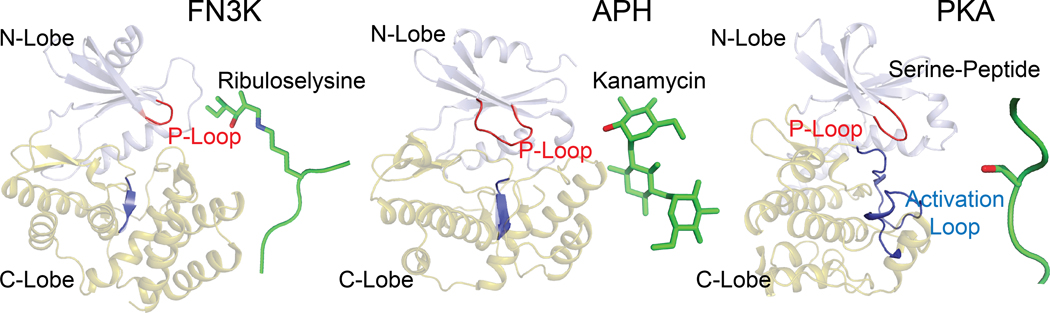
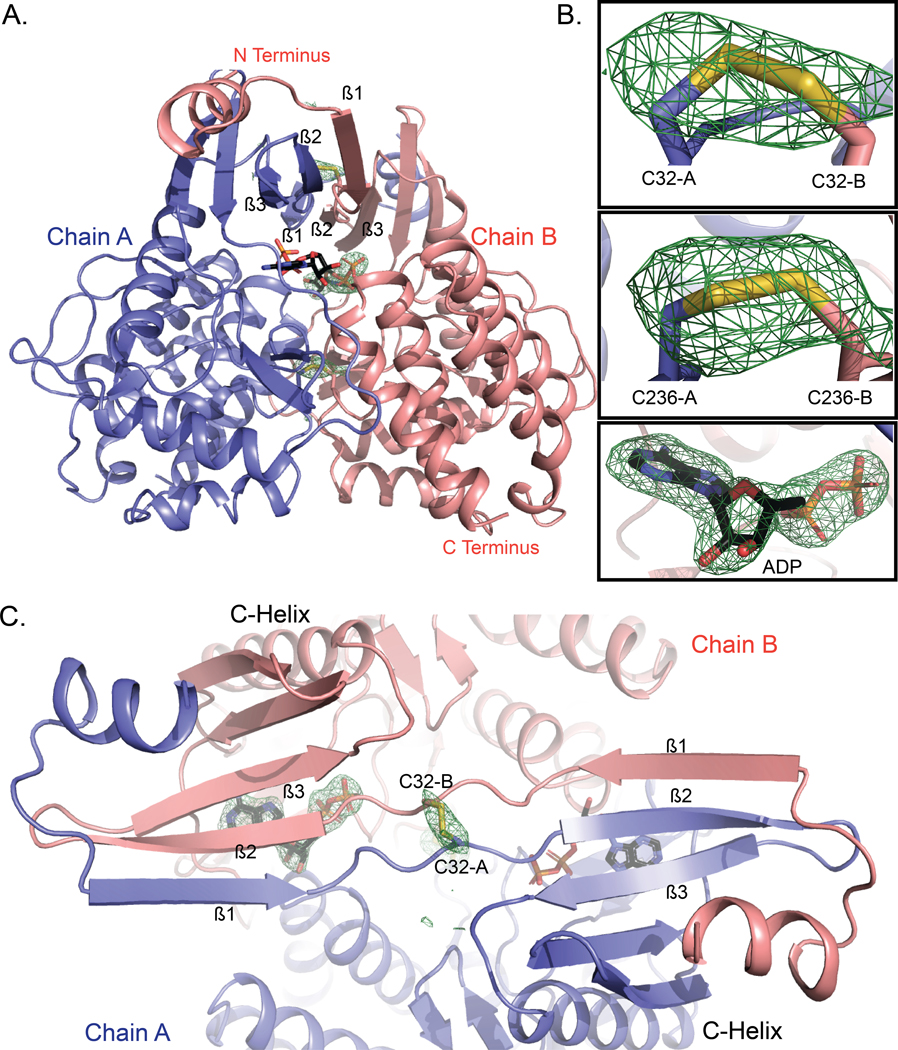
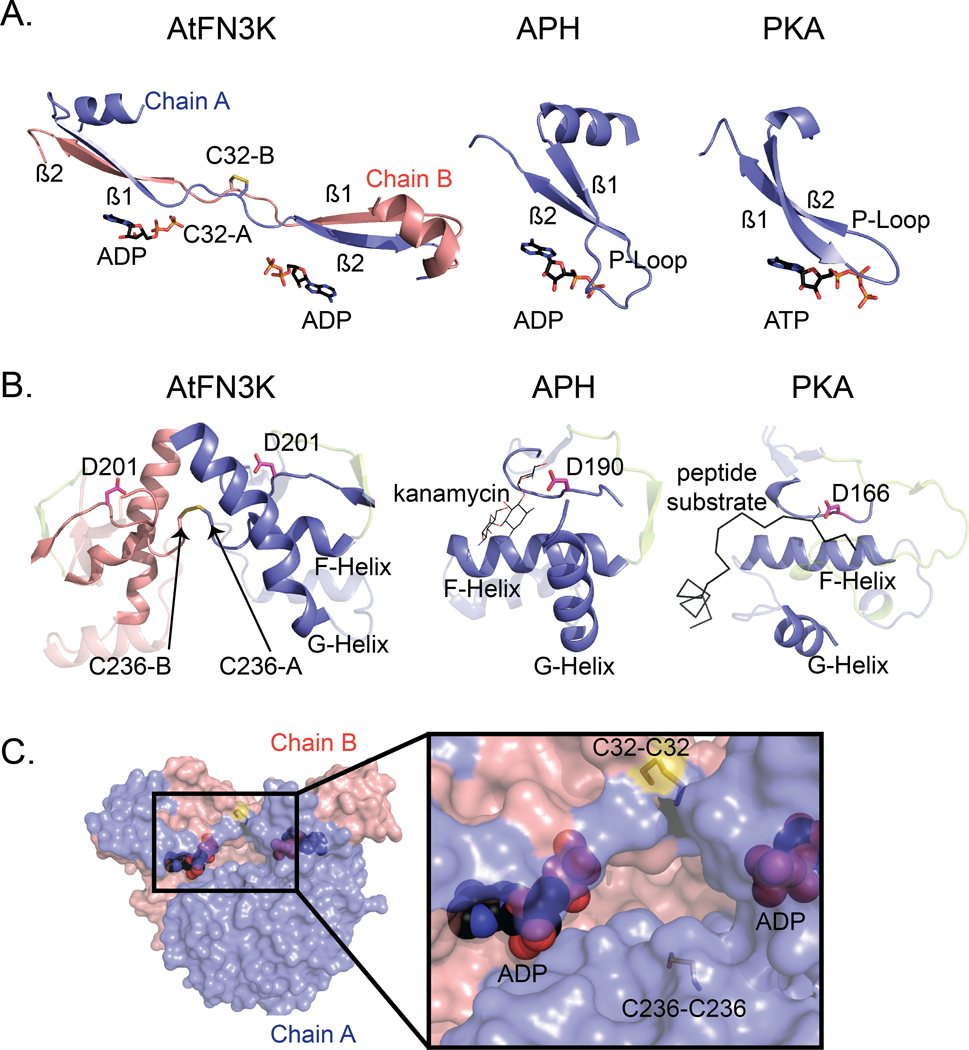
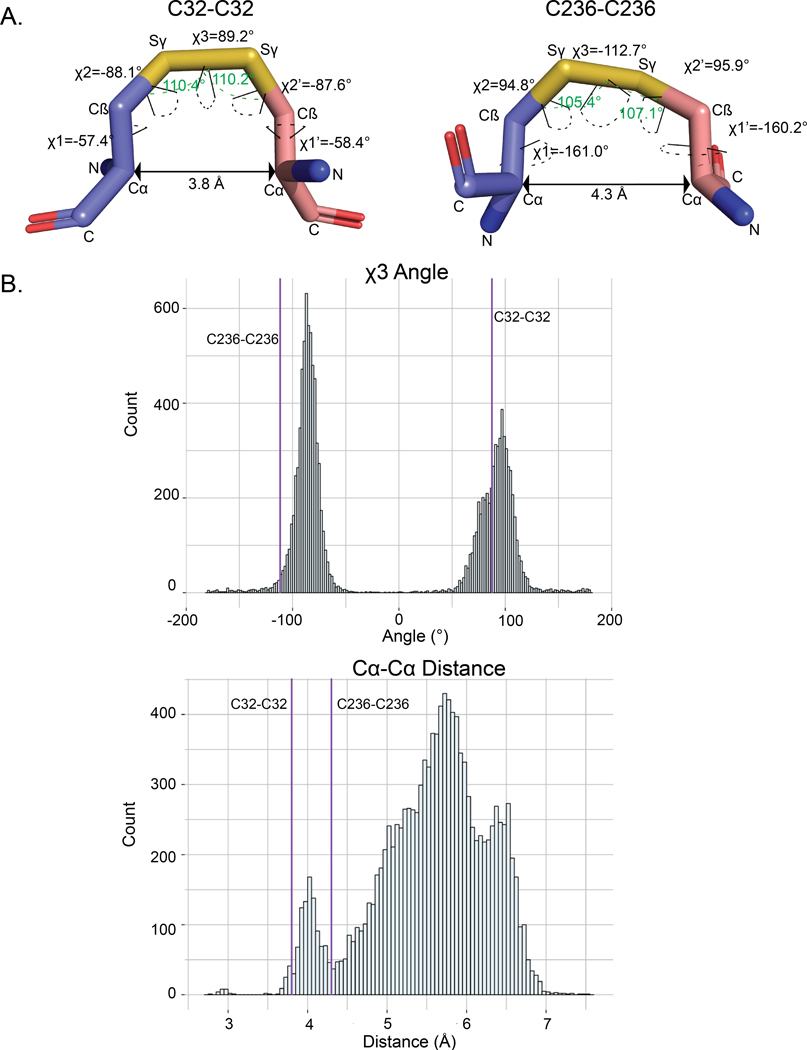
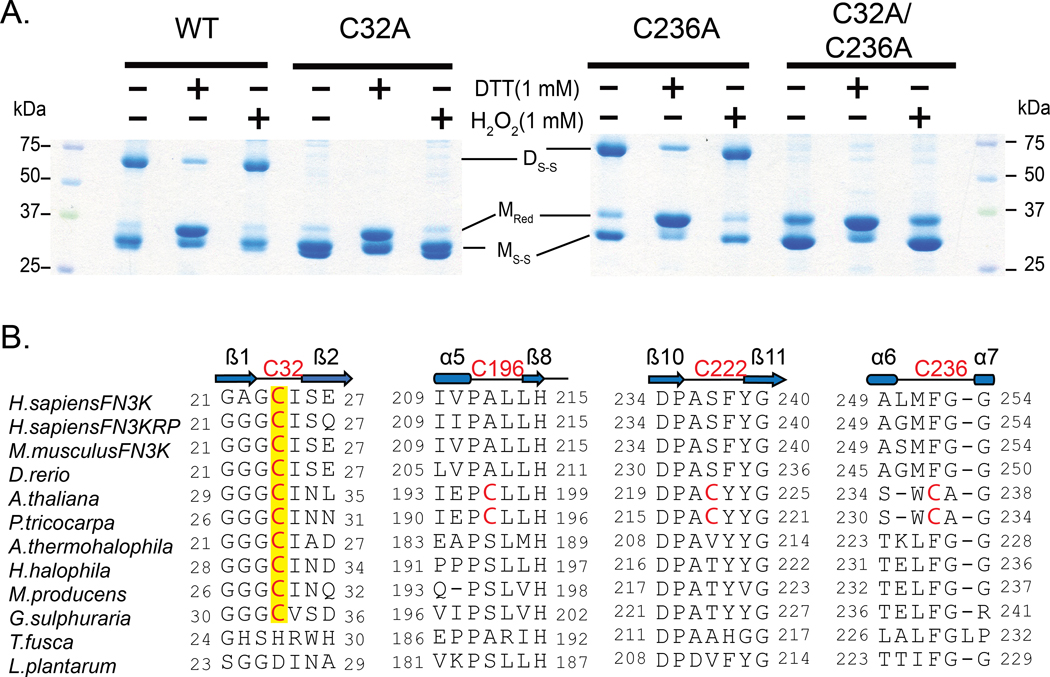
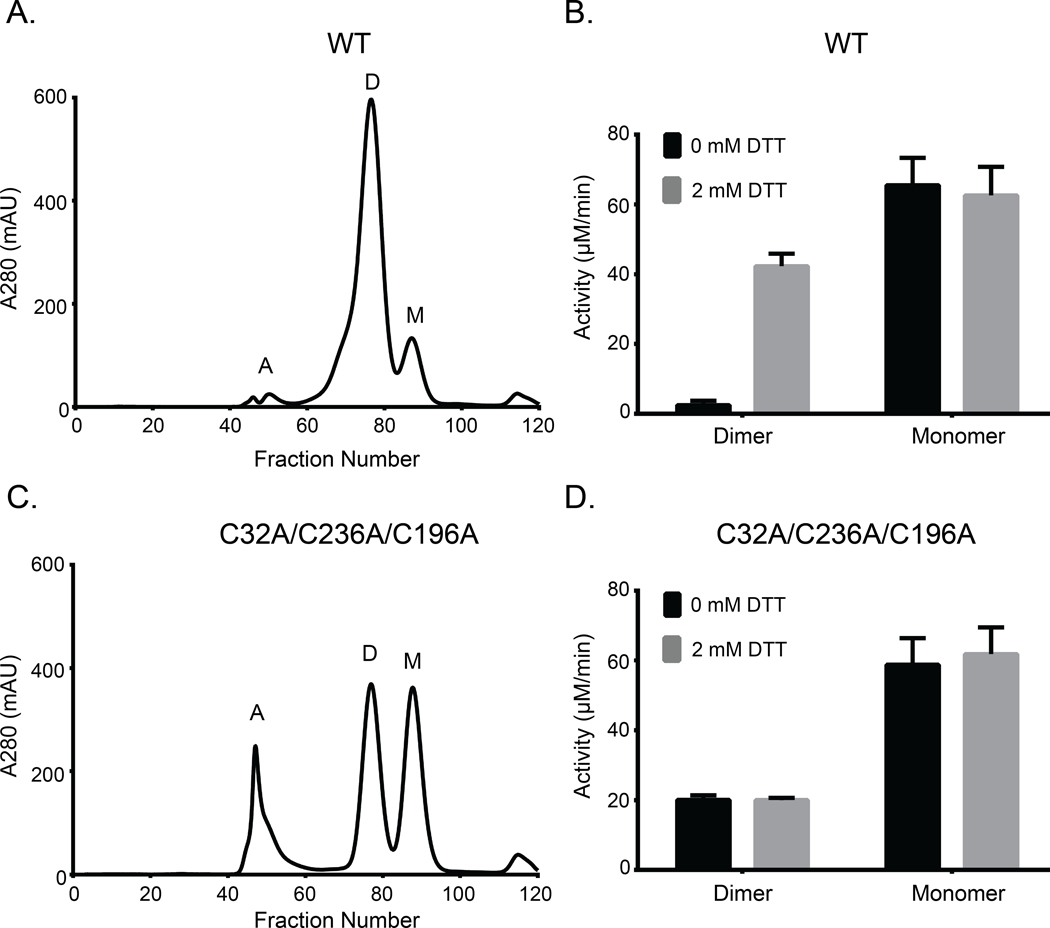
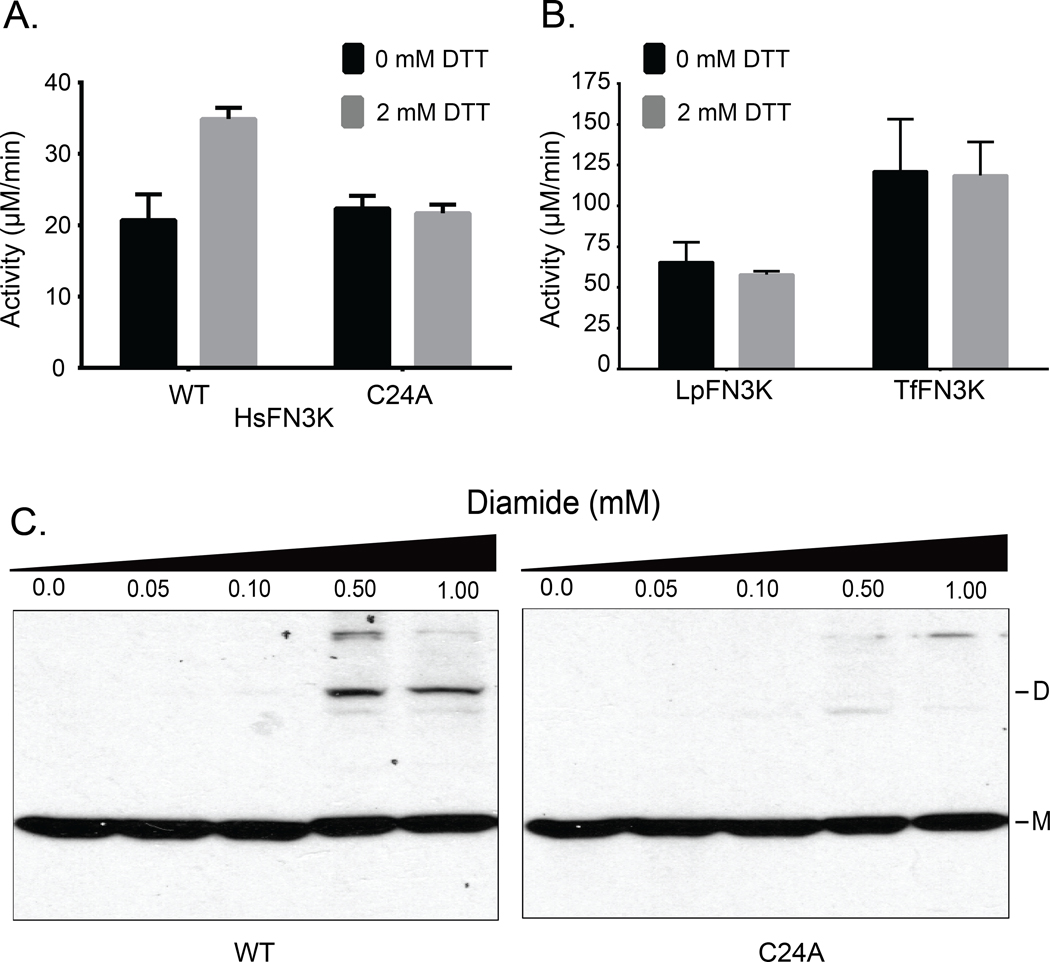
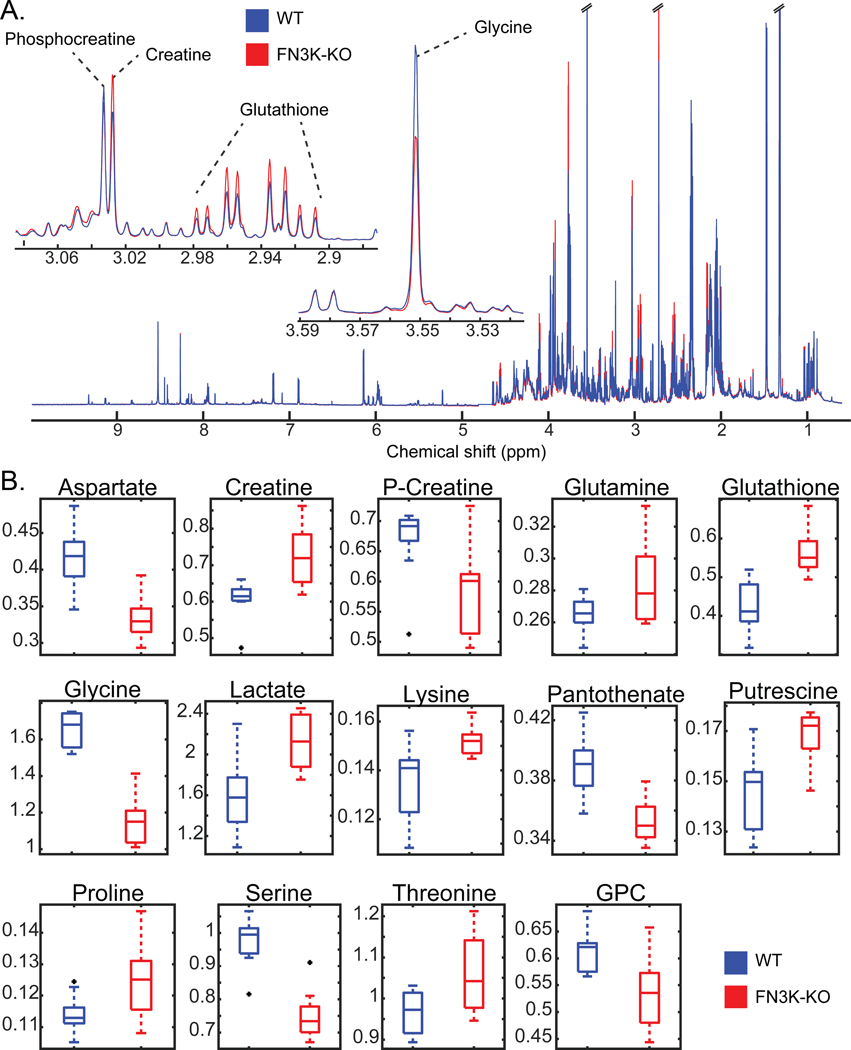
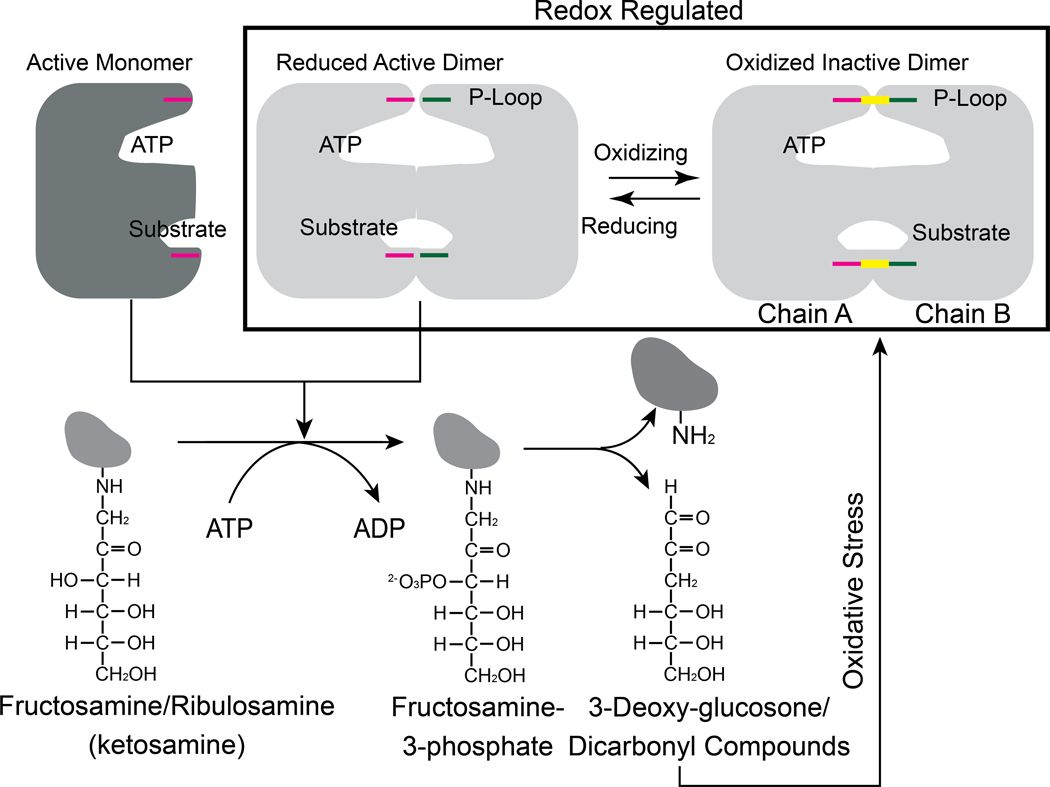
Aberrant regulation of metabolic kinases by altered redox homeostasis substantially contributes to aging and various diseases, such as diabetes. We found that the catalytic activity of a conserved family of fructosamine-3-kinases (FN3Ks), which are evolutionarily related to eukaryotic protein kinases, is regulated by redox-sensitive cysteine residues in the kinase domain. The crystal structure of the FN3K homolog from Arabidopsis thaliana revealed that it forms an unexpected strand-exchange dimer in which the ATP-binding P-loop and adjoining β strands are swapped between two chains in the dimer. This dimeric configuration is characterized by strained interchain disulfide bonds that stabilize the P-loop in an extended conformation. Mutational analysis and solution studies confirmed that the strained disulfides function as redox "switches" to reversibly regulate the activity and dimerization of FN3K. Human FN3K, which contains an equivalent P-loop Cys, was also redox sensitive, whereas ancestral bacterial FN3K homologs, which lack a P-loop Cys, were not. Furthermore, CRISPR-mediated knockout of FN3K in human liver cancer cells altered the abundance of redox metabolites, including an increase in glutathione. We propose that redox regulation evolved in FN3K homologs in response to changing cellular redox conditions. Our findings provide insights into the origin and evolution of redox regulation in the protein kinase superfamily and may open new avenues for targeting human FN3K in diabetic complications.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures
References
-
- Maillard LC, The action of amino acids on sugar; the formation of melanoidin by a methodic route. Comptes Ren- dus Hebdomadaires Des Seances De L Academie Des Sci- ences 154, 66–68 (1912).
-
- Hodge JE, The Amadori rearrangement. Adv Carbohydr Chem 10, 169–205 (1955). - PubMed
-
- Higgins H. F. B. a. P. J., Reaction of Monosaccharides with Proteins: Possible Evolutionary Significance. Science 213, 222–224 (1981). - PubMed
-
- Baynes JW, Watkins NG, Fisher CI, Hull CJ, Patrick JS, Ahmed MU, Dunn JA, Thorpe SR, The Amadori product on protein: structure and reactions. Prog Clin Biol Res 304, 43–67 (1989). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
