

Challenges and advances in genome mining of ribosomally synthesized and post-translationally modified peptides (RiPPs)
- PMID: 32637669
- PMCID: PMC7327761
- DOI: 10.1016/j.synbio.2020.06.002
Challenges and advances in genome mining of ribosomally synthesized and post-translationally modified peptides (RiPPs)
Erratum in
-
Erratum regarding previously published articles.Synth Syst Biotechnol. 2020 Oct 14;5(4):330-331. doi: 10.1016/j.synbio.2020.10.001. eCollection 2020 Dec. Synth Syst Biotechnol. 2020. PMID: 33102827 Free PMC article.
Abstract
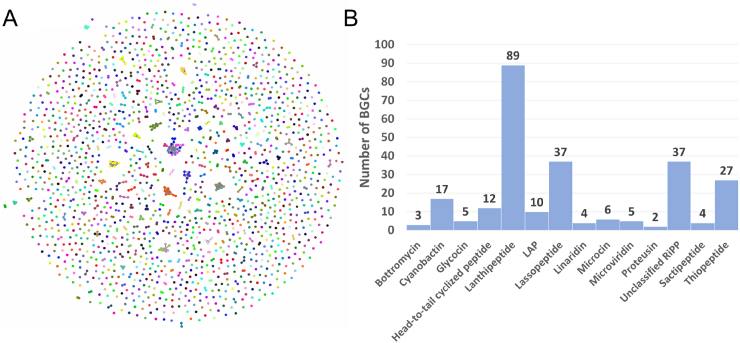
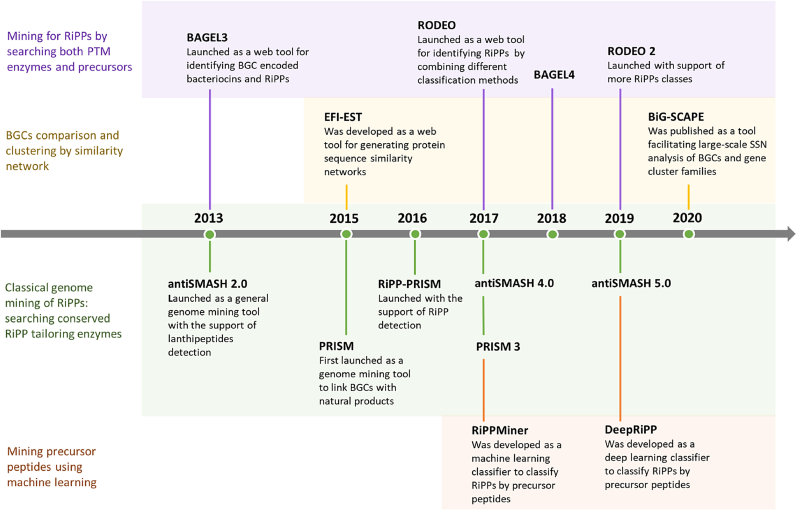
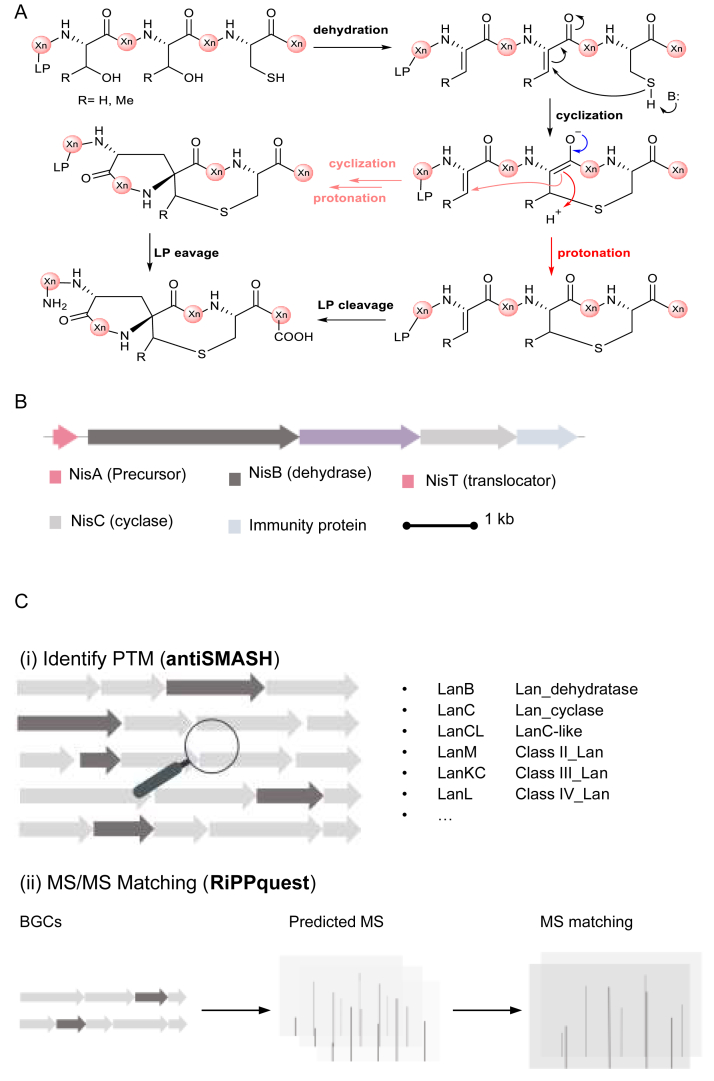
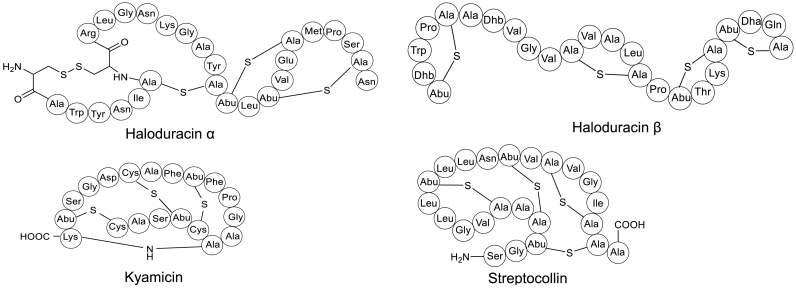
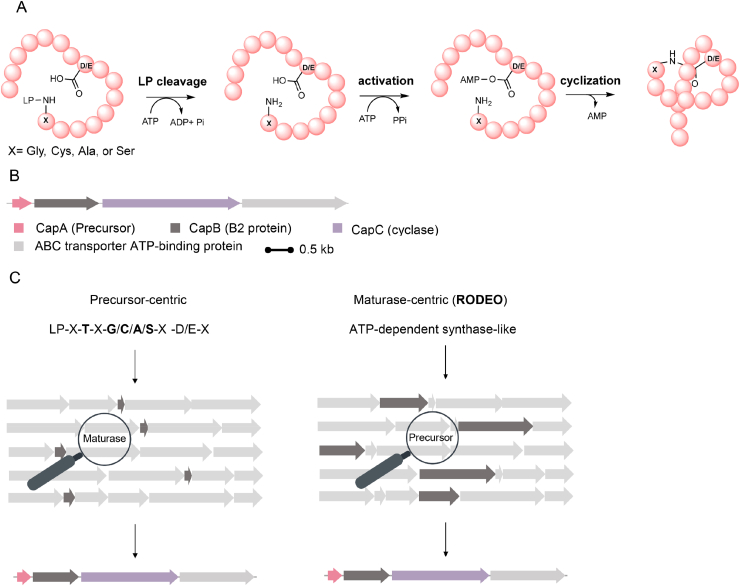
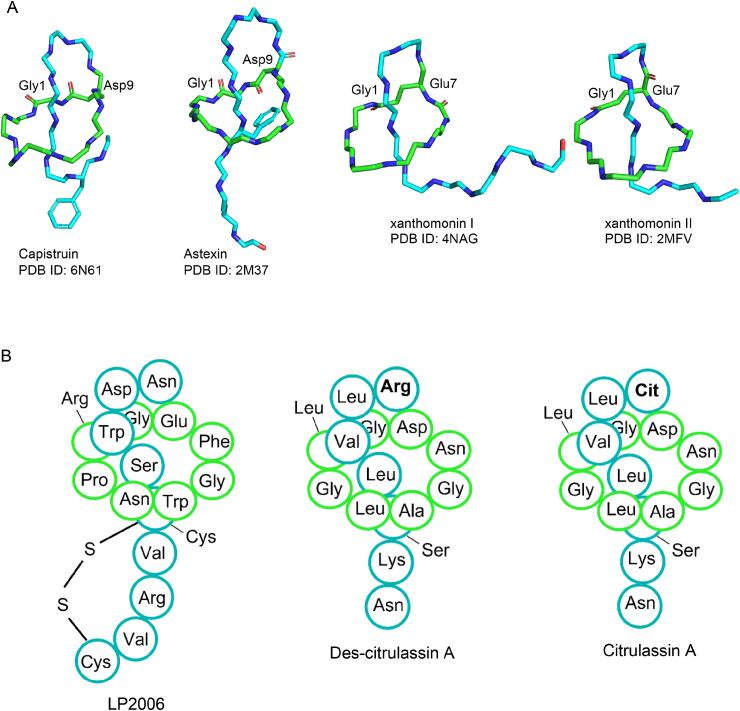
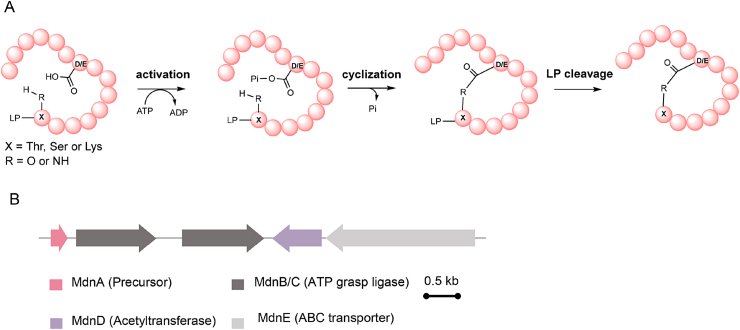
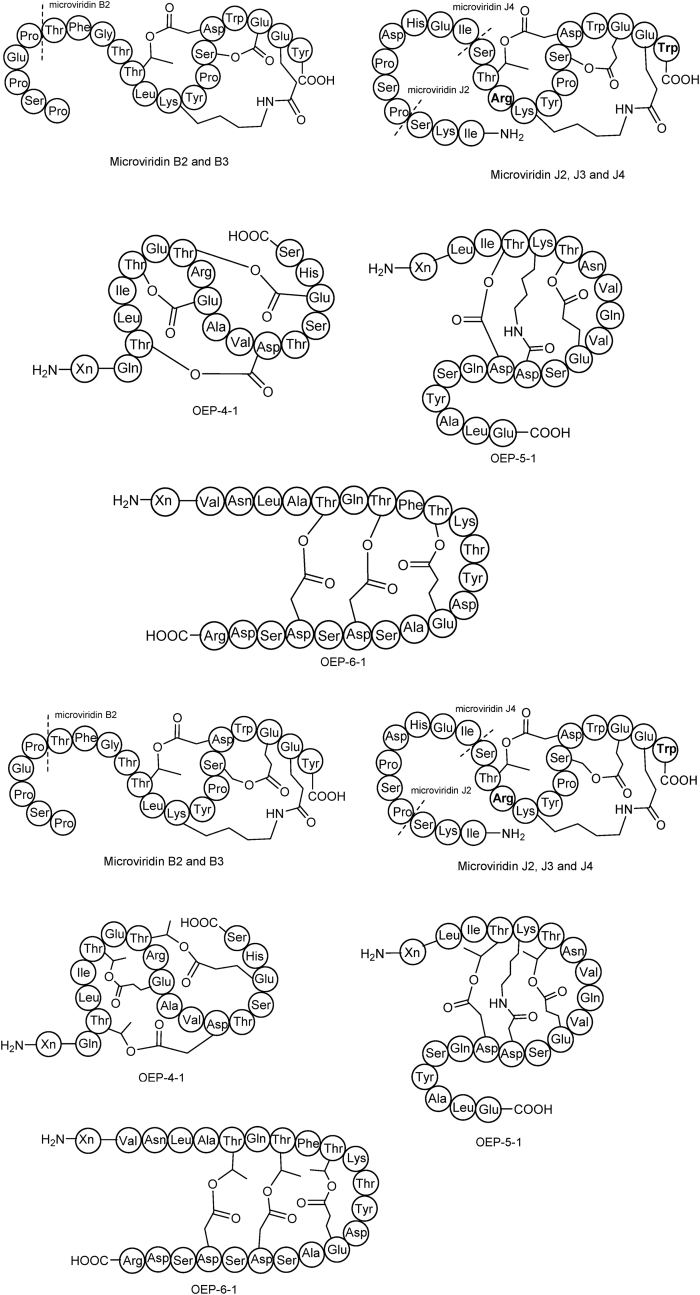
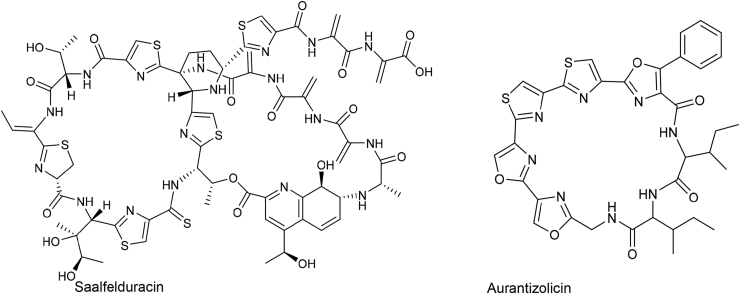
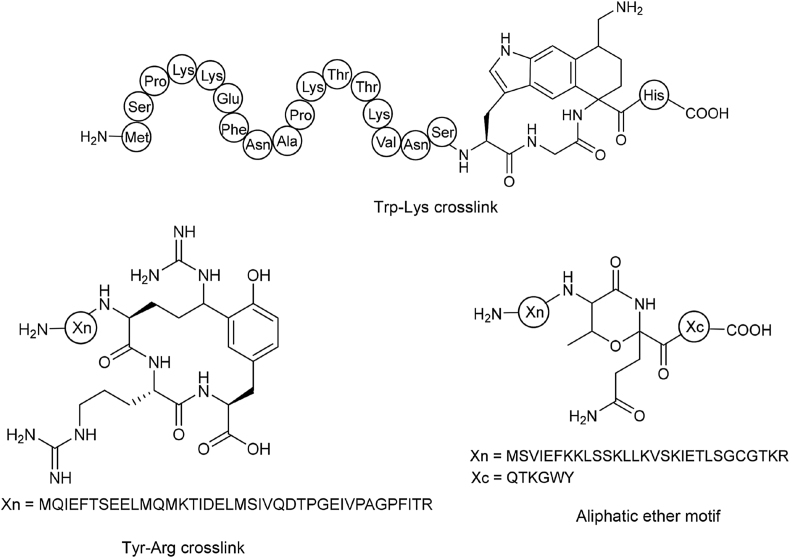
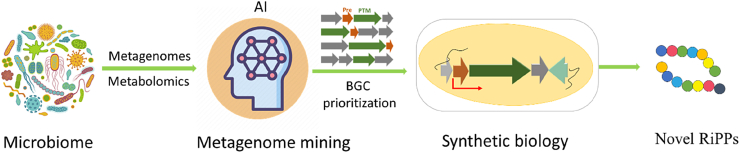
Ribosomally synthesized and post-translationally modified peptides (RiPPs) are a class of cyclic or linear peptidic natural products with remarkable structural and functional diversity. Recent advances in genomics and synthetic biology, are facilitating us to discover a large number of new ribosomal natural products, including lanthipeptides, lasso peptides, sactipeptides, thiopeptides, microviridins, cyanobactins, linear thiazole/oxazole-containing peptides and so on. In this review, we summarize bioinformatic strategies that have been developed to identify and prioritize biosynthetic gene clusters (BGCs) encoding RiPPs, and the genome mining-guided discovery of novel RiPPs. We also prospectively provide a vision of what genomics-guided discovery of RiPPs may look like in the future, especially the discovery of RiPPs from dominant but uncultivated microbes, which will be promoted by the combinational use of synthetic biology and metagenome mining strategies.
Keywords: Metagenome mining; Natural products; RiPPs, Genome mining; Ribosomally synthesized and post-translationally modified peptides; Synthetic biology.
© 2020 KeAi Communications Co.(+) Ltd.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
