

This is a preprint.
Mutations from bat ACE2 orthologs markedly enhance ACE2-Fc neutralization of SARS-CoV-2
- PMID: 32637954
- PMCID: PMC7337384
- DOI: 10.1101/2020.06.29.178459
Mutations from bat ACE2 orthologs markedly enhance ACE2-Fc neutralization of SARS-CoV-2
Update in
-
Mutations derived from horseshoe bat ACE2 orthologs enhance ACE2-Fc neutralization of SARS-CoV-2.PLoS Pathog. 2021 Apr 9;17(4):e1009501. doi: 10.1371/journal.ppat.1009501. eCollection 2021 Apr. PLoS Pathog. 2021. PMID: 33836016 Free PMC article.
Abstract
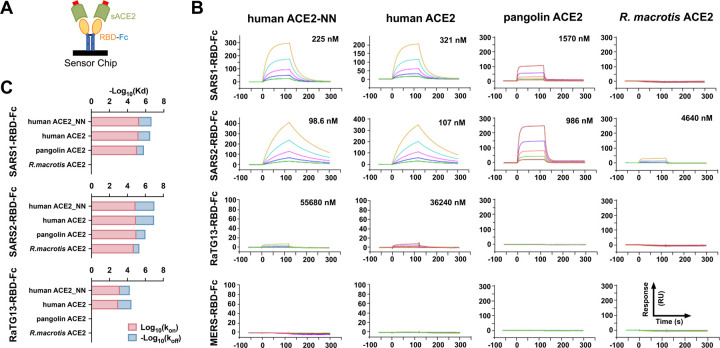
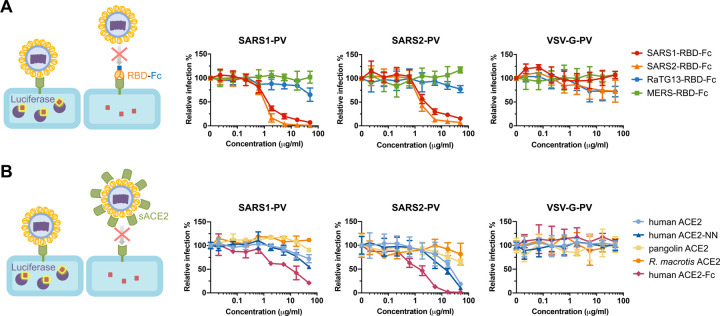
The severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) spike (S) protein mediates infection of cells expressing angiotensin-converting enzyme 2 (ACE2). ACE2 is also the viral receptor of SARS-CoV (SARS-CoV-1), a related coronavirus that emerged in 2002-2003. Horseshoe bats (genus Rhinolophus ) are presumed to be the original reservoir of both viruses, and a SARS-like coronavirus, RaTG13, closely related SARS-CoV-2, has been isolated from one horseshoe-bat species. Here we characterize the ability of S-protein receptor-binding domains (RBDs) of SARS-CoV-1, SARS-CoV-2, and RaTG13 to bind a range of ACE2 orthologs. We observed that the SARS-CoV-2 RBD bound human, pangolin, and horseshoe bat ( R. macrotis) ACE2 more efficiently than the SARS-CoV-1 or RaTG13 RBD. Only the RaTG13 RBD bound rodent ACE2 orthologs efficiently. Five mutations drawn from ACE2 orthologs of nine Rhinolophus species enhanced human ACE2 binding to the SARS-CoV-2 RBD and neutralization of SARS-CoV-2 by an immunoadhesin form of human ACE2 (ACE2-Fc). Two of these mutations impaired neutralization of SARS-CoV-1. An ACE2-Fc variant bearing all five mutations neutralized SARS-CoV-2 five-fold more efficiently than human ACE2-Fc. These data narrow the potential SARS-CoV-2 reservoir, suggest that SARS-CoV-1 and -2 originate from distinct bat species, and identify a more potently neutralizing form of ACE2-Fc.
Conflict of interest statement
Declaration of Interests
M.R.G., C.C.B., M.D.A., and M.F. are all cofounders of, and have an equity interest in Emmune Inc., a small biotech company that specializes in the development of antibody-like antiviral therapies.
Figures

References
-
- Guan Y., Zheng B.J., He Y.Q., Liu X.L., Zhuang Z.X., Cheung C.L., Luo S.W., Li P.H., Zhang L.J., Guan Y.J., et al. (2003). Isolation and characterization of viruses related to the SARS coronavirus from animals in southern China. Science 302, 276–278. - PubMed
-
- Lan J., Ge J., Yu J., Shan S., Zhou H., Fan S., Zhang Q., Shi X., Wang Q., Zhang L., et al. (2020). Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 581, 215–220. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous