

COVID-19 and the Chemical Senses: Supporting Players Take Center Stage
- PMID: 32640192
- PMCID: PMC7328585
- DOI: 10.1016/j.neuron.2020.06.032
COVID-19 and the Chemical Senses: Supporting Players Take Center Stage
Abstract
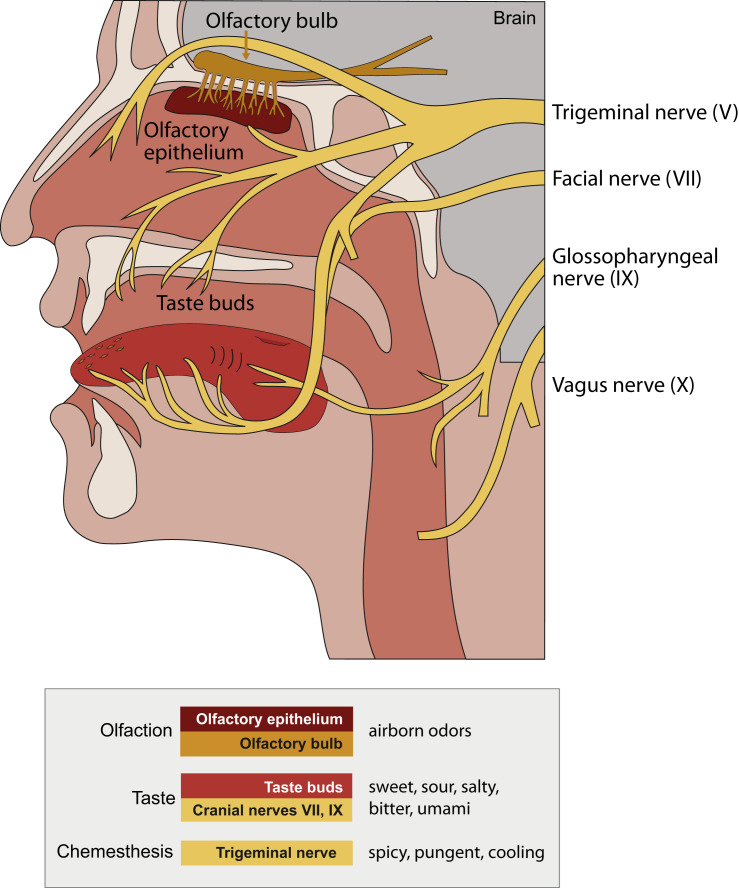
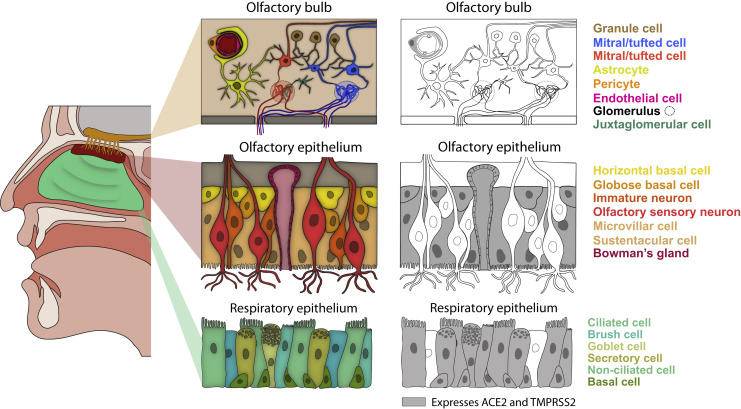
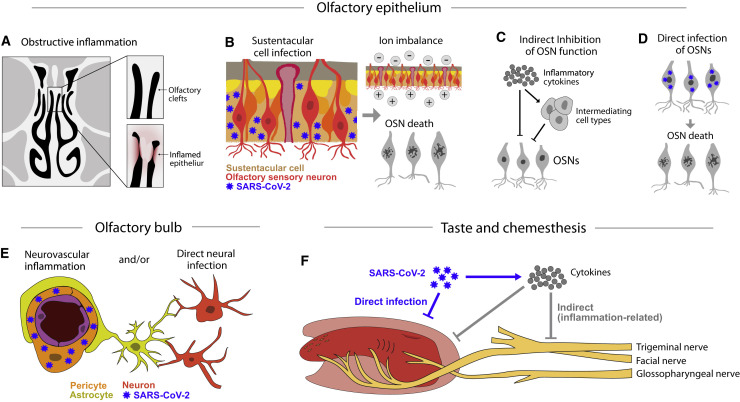
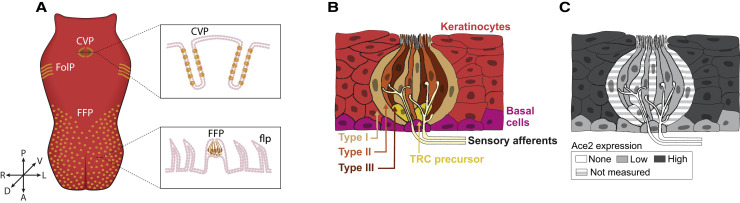
The main neurological manifestation of COVID-19 is loss of smell or taste. The high incidence of smell loss without significant rhinorrhea or nasal congestion suggests that SARS-CoV-2 targets the chemical senses through mechanisms distinct from those used by endemic coronaviruses or other common cold-causing agents. Here we review recently developed hypotheses about how SARS-CoV-2 might alter the cells and circuits involved in chemosensory processing and thereby change perception. Given our limited understanding of SARS-CoV-2 pathogenesis, we propose future experiments to elucidate disease mechanisms and highlight the relevance of this ongoing work to understanding how the virus might alter brain function more broadly.
Copyright © 2020 Elsevier Inc. All rights reserved.
Figures
References
-
- Adamczyk K., Herman M., Fraczek J., Piec R., Szykula-Piec B., Zaczynski A., Wojtowicz R., Bojanowski K., Rusyan E., Krol Z., et al. Sensitivity and specifity of prediction models based on gustatory disorders in diagnosing COVID-19 patients: a case-control study. medRxiv. 2020 doi: 10.1101/2020.05.31.20118380. - DOI
-
- Åkerlund A., Bende M., Murphy C. Olfactory threshold and nasal mucosal changes in experimentally induced common cold. Acta Otolaryngol. 1995;115:88–92. - PubMed
-
- Ansari M.A., Marchi E., Ramamurthy N., Aschenbrenner D., Hackstein C.-P., Bowden R., Sharma E., Pedergnana V., Venkateswaran S., Kugathasan S., et al. A gene locus that controls expression of ACE2 in virus infection. medRxiv. 2020 doi: 10.1101/2020.04.26.20080408. - DOI
-
- Armulik A., Genové G., Betsholtz C. Pericytes: developmental, physiological, and pathological perspectives, problems, and promises. Dev. Cell. 2011;21:193–215. - PubMed
-
- Bader M. Tissue renin-angiotensin-aldosterone systems: Targets for pharmacological therapy. Annu. Rev. Pharmacol. Toxicol. 2010;50:439–465. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
