

Envelope protein ubiquitination drives entry and pathogenesis of Zika virus
- PMID: 32641828
- PMCID: PMC7501154
- DOI: 10.1038/s41586-020-2457-8
Envelope protein ubiquitination drives entry and pathogenesis of Zika virus
Abstract
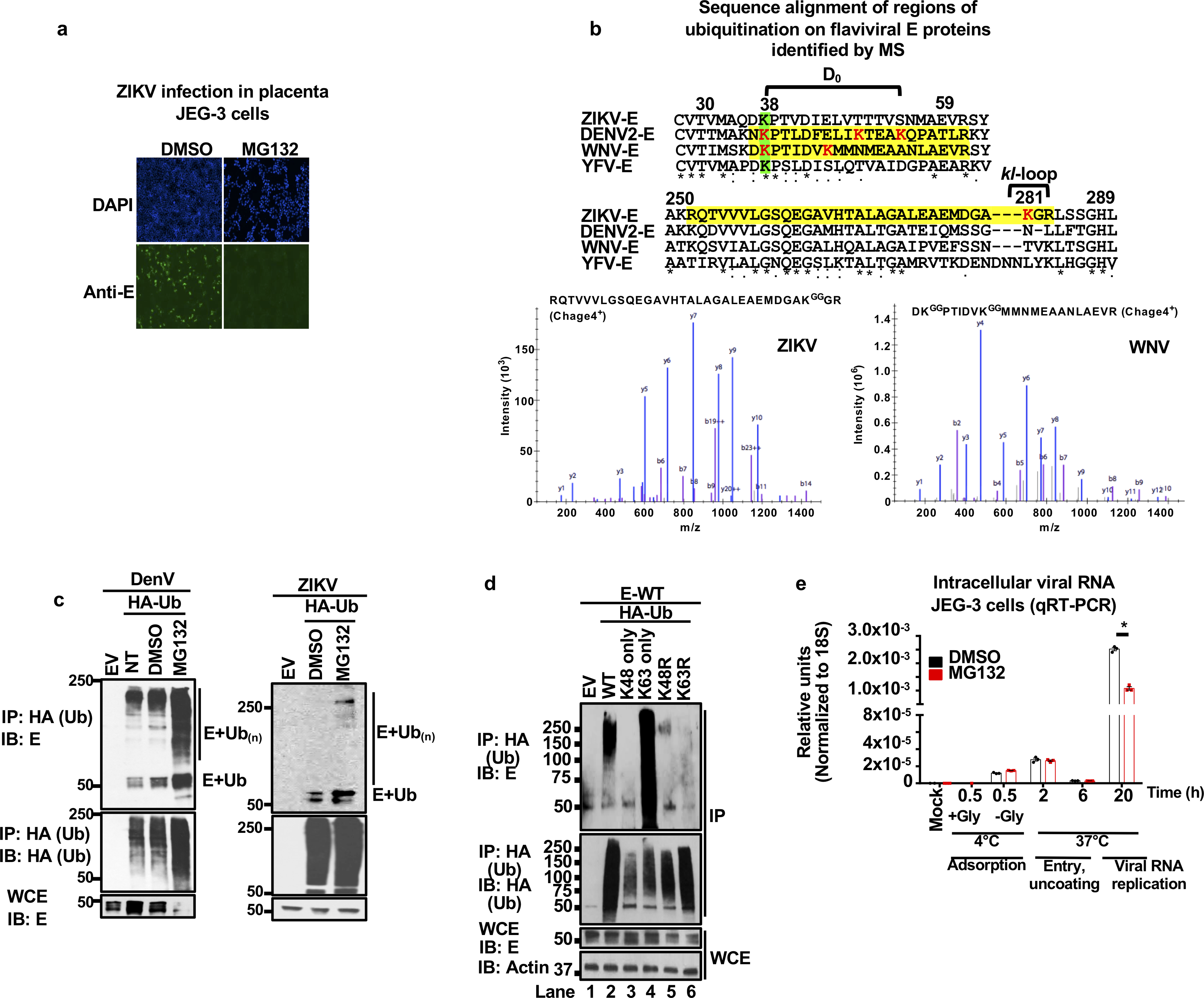
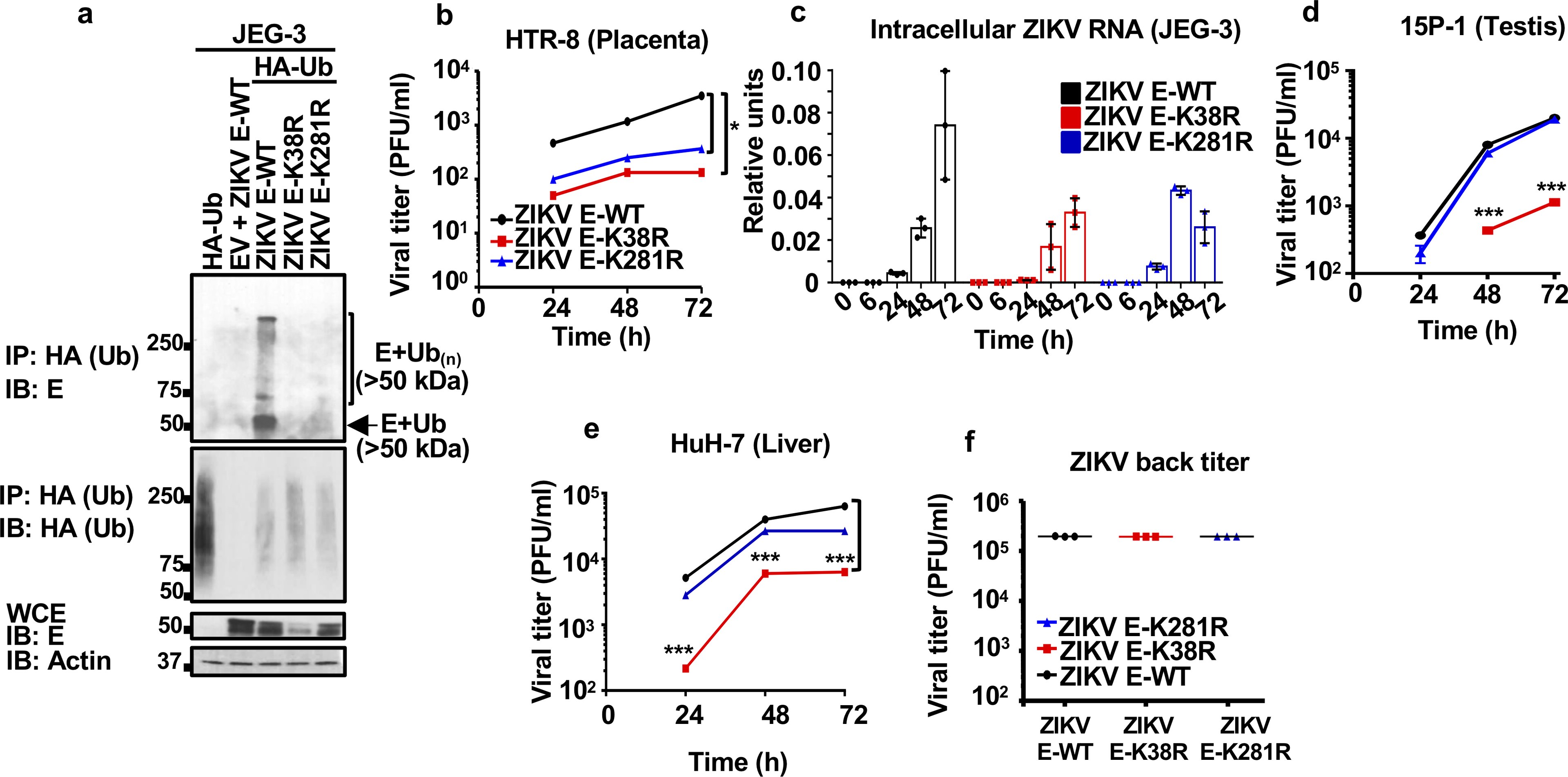
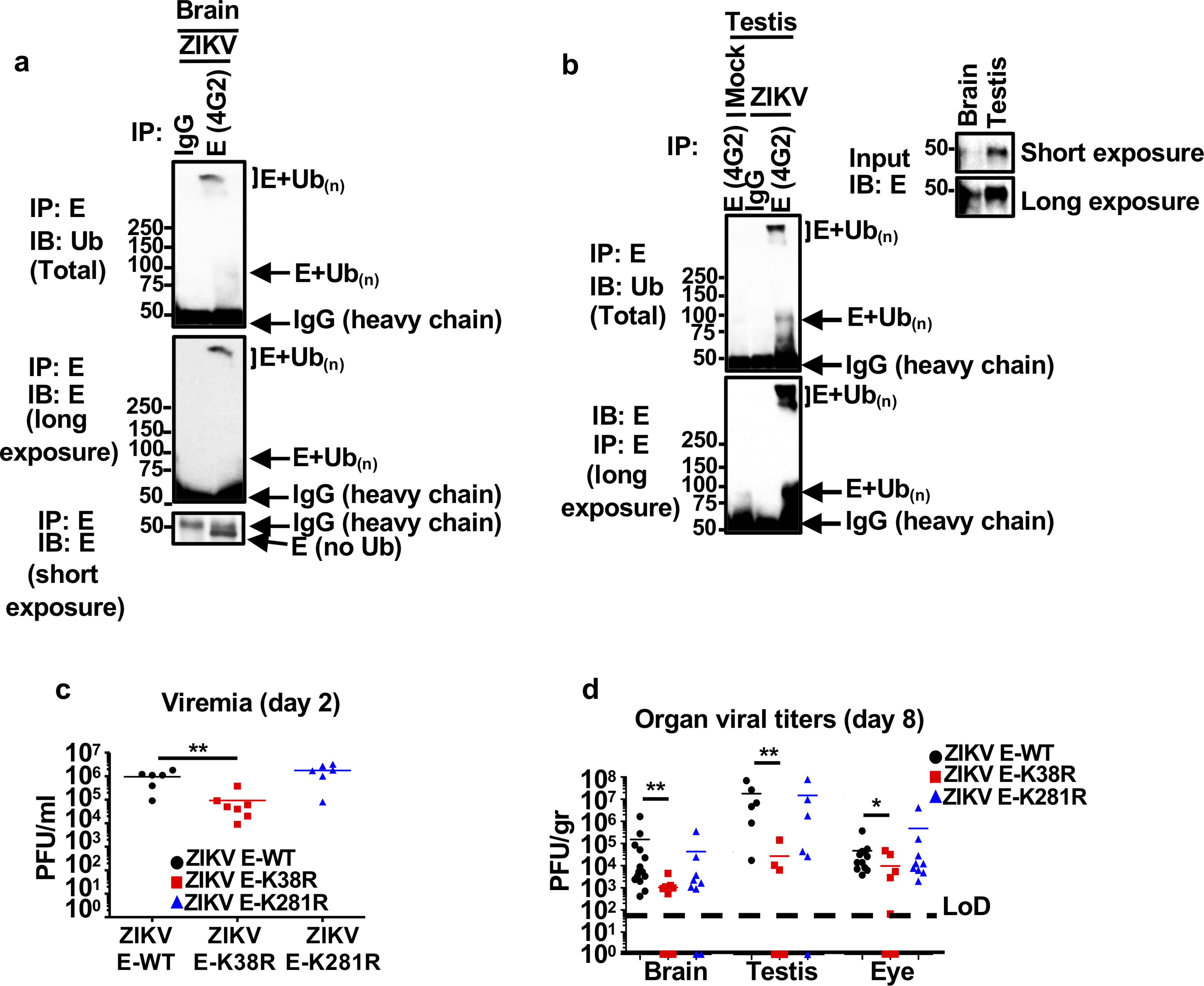
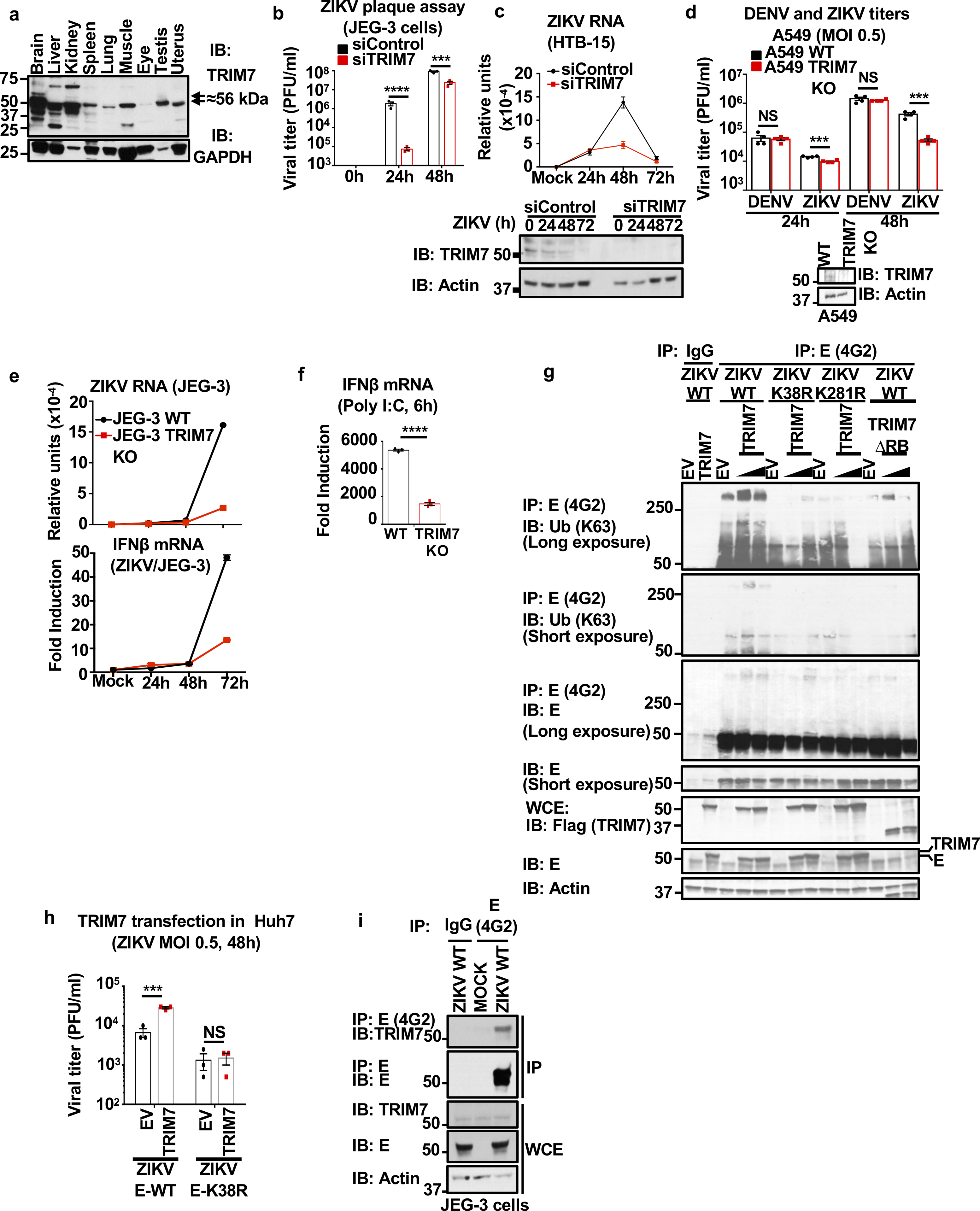
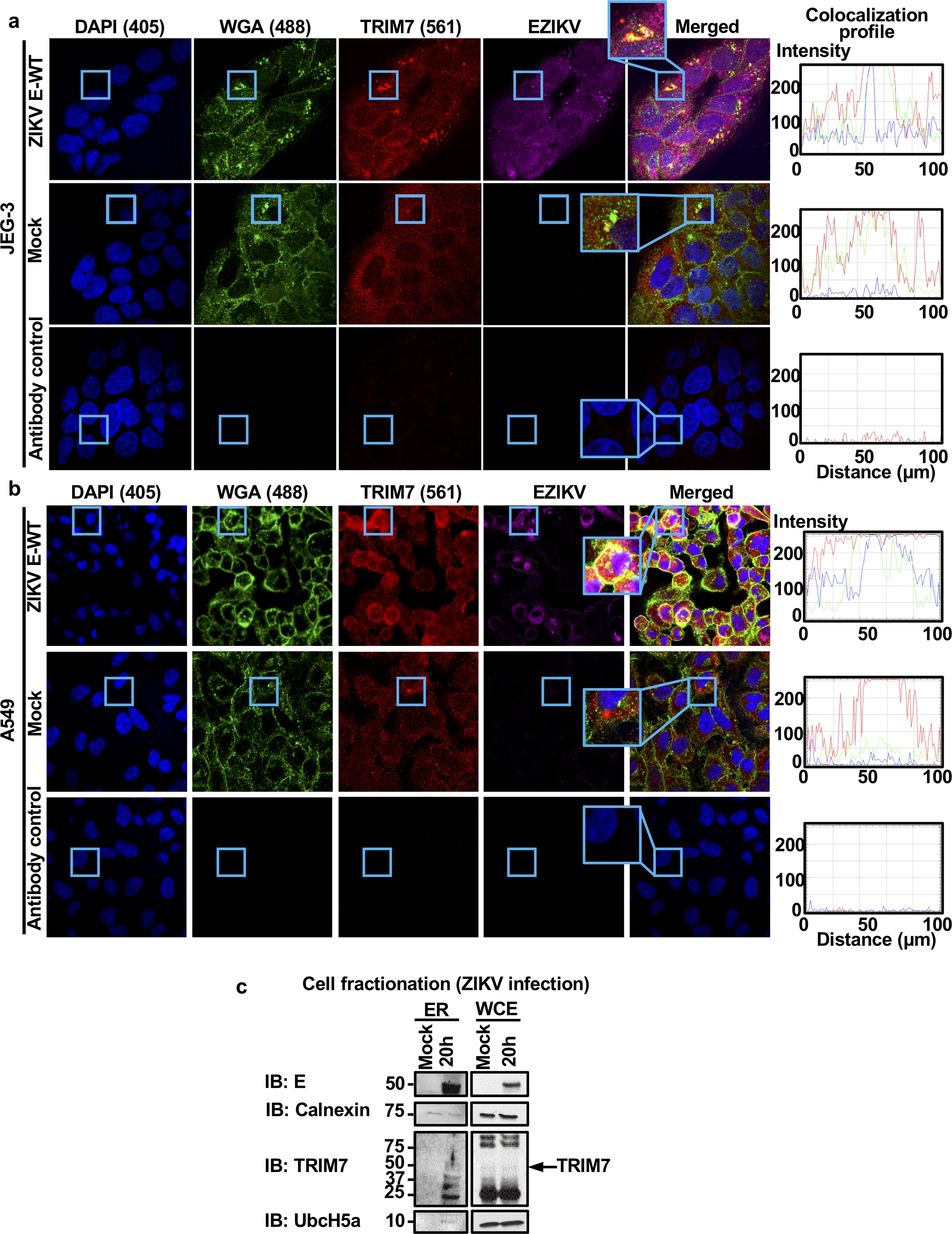
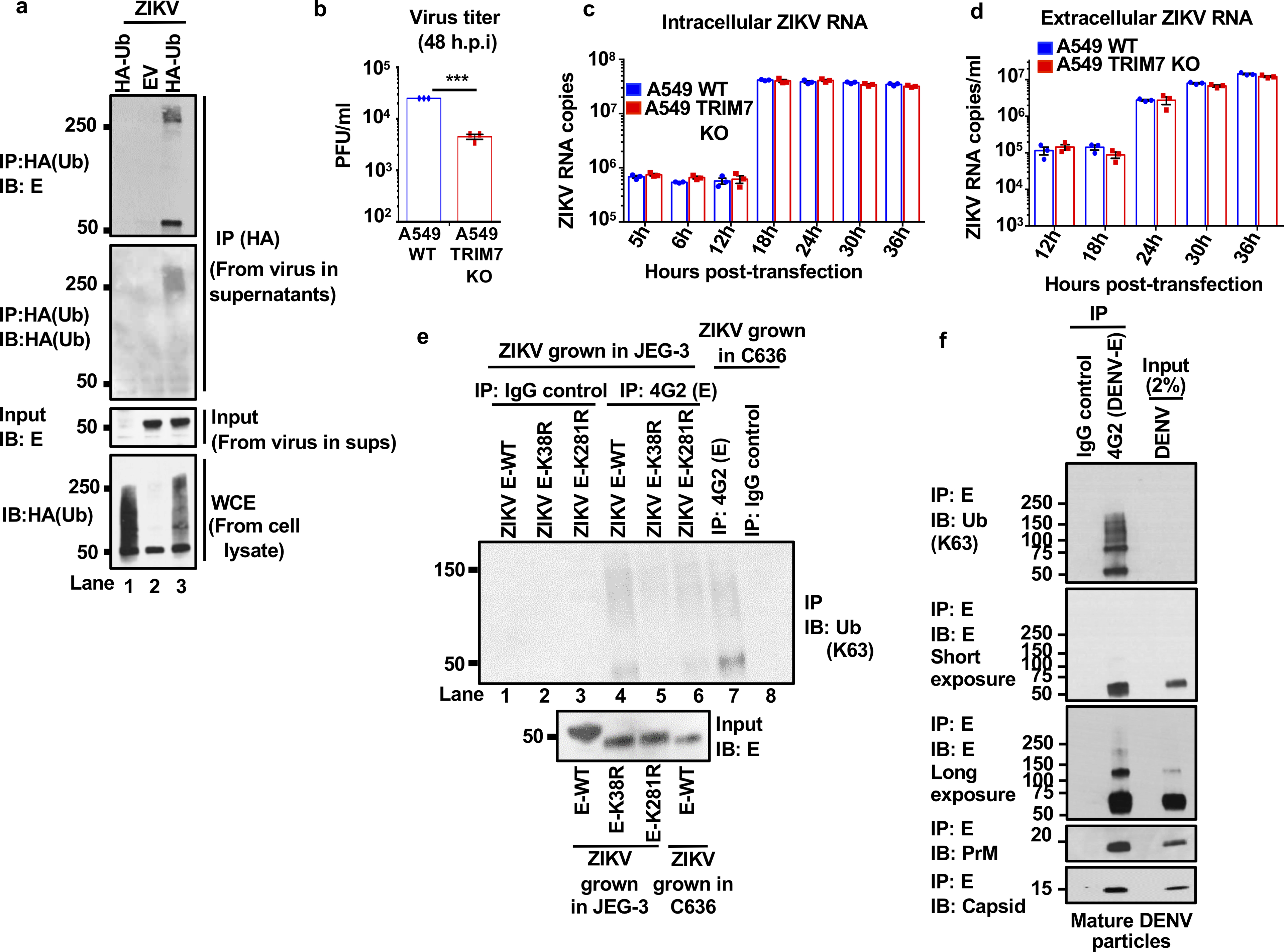
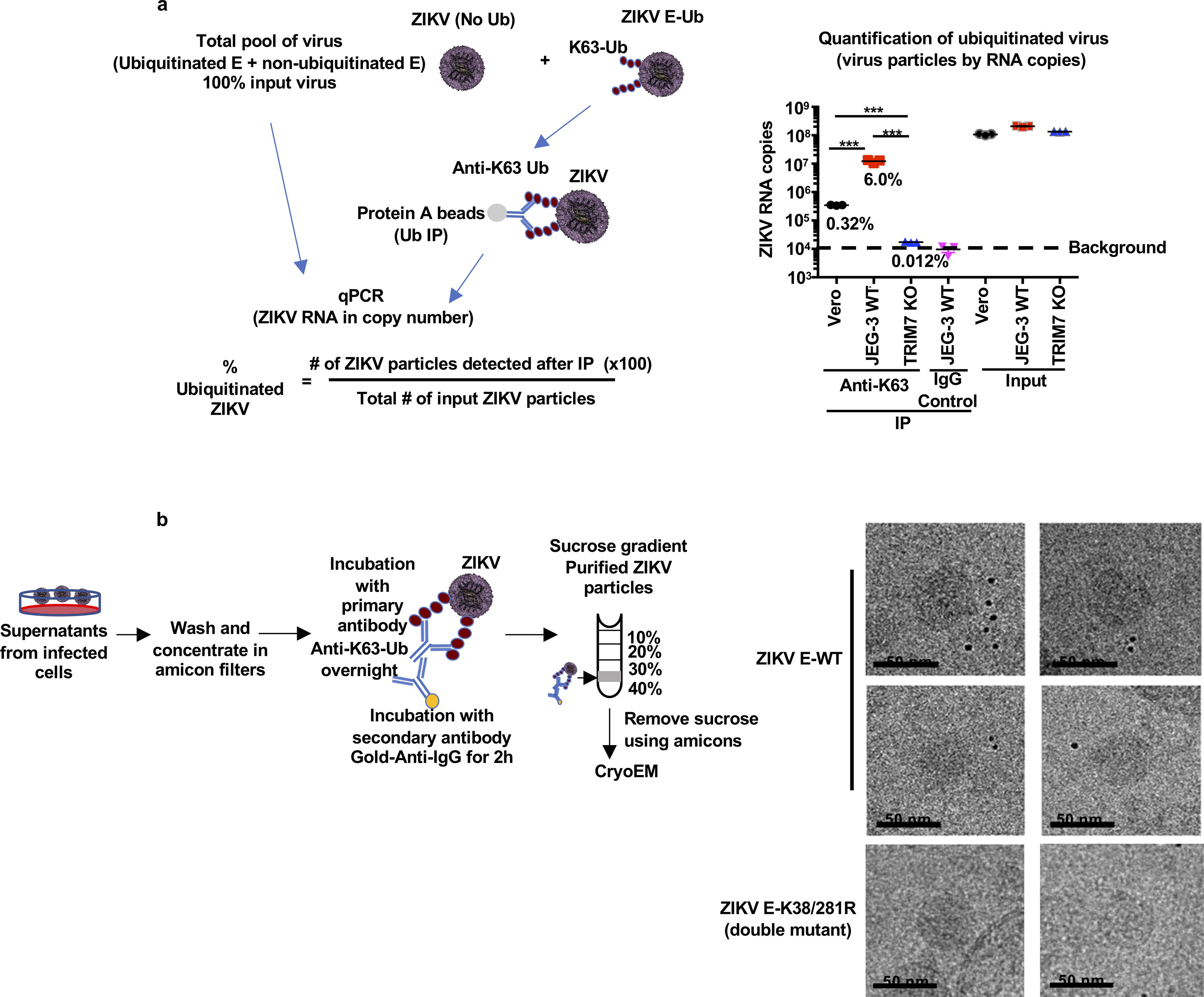
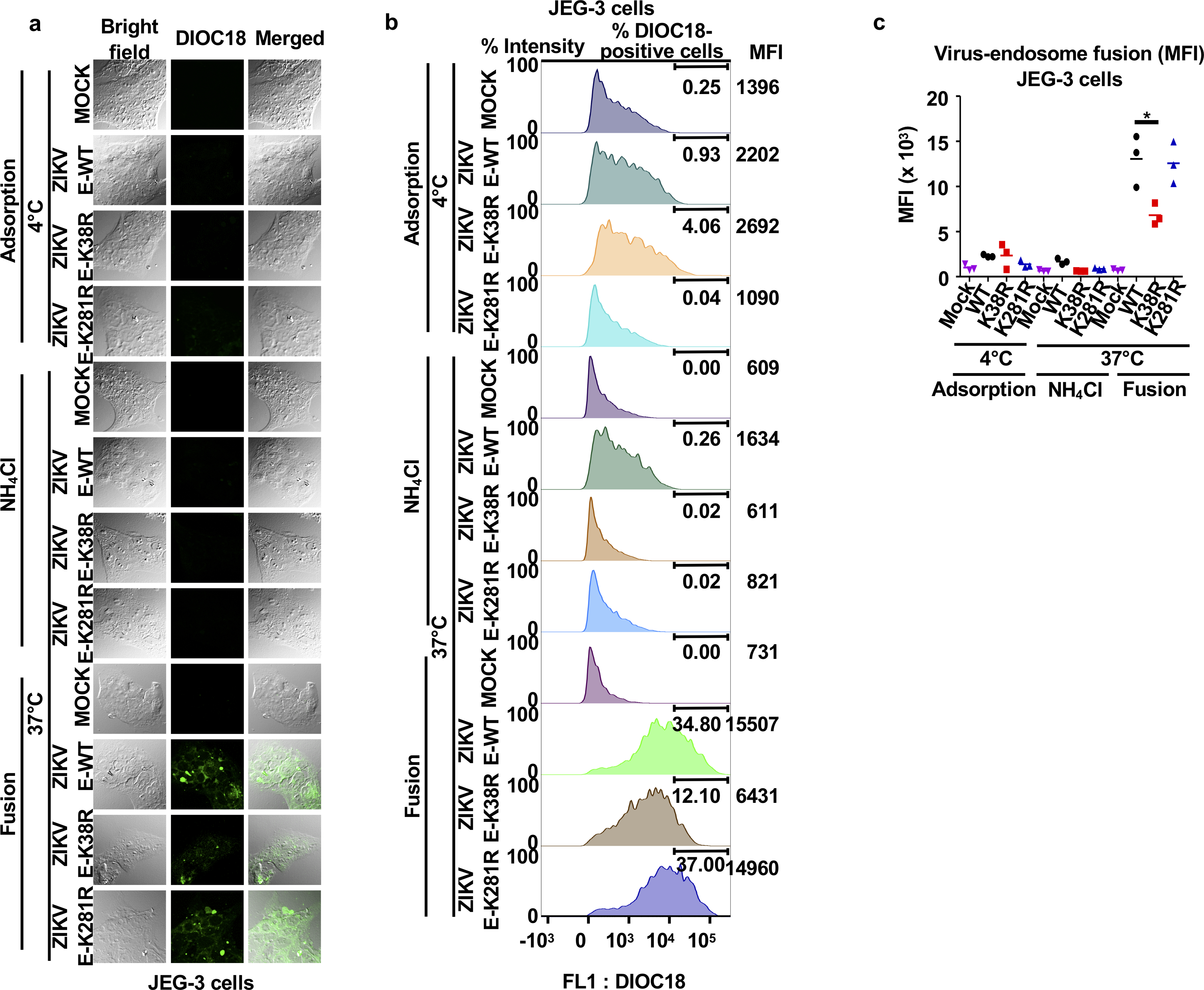
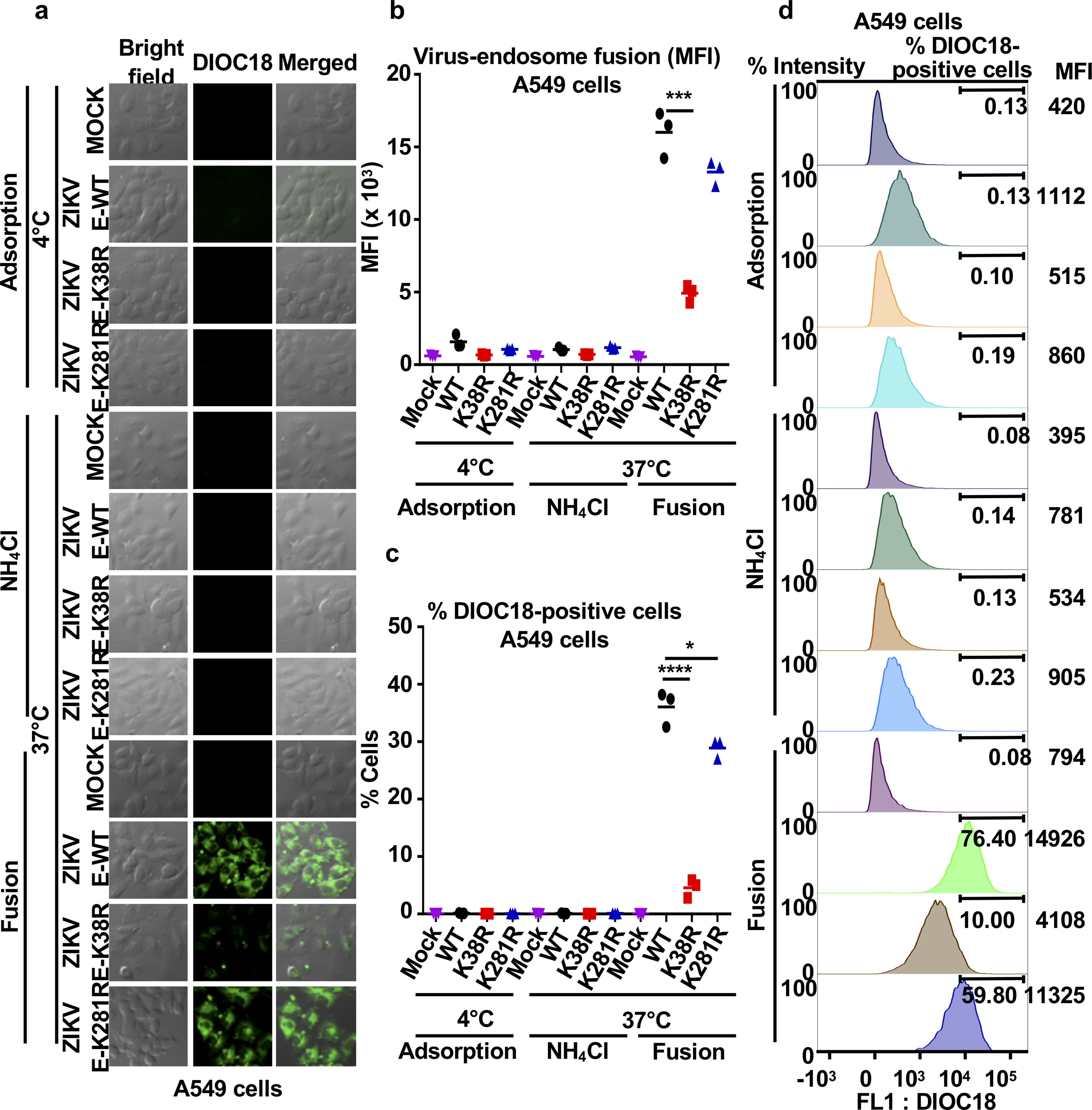
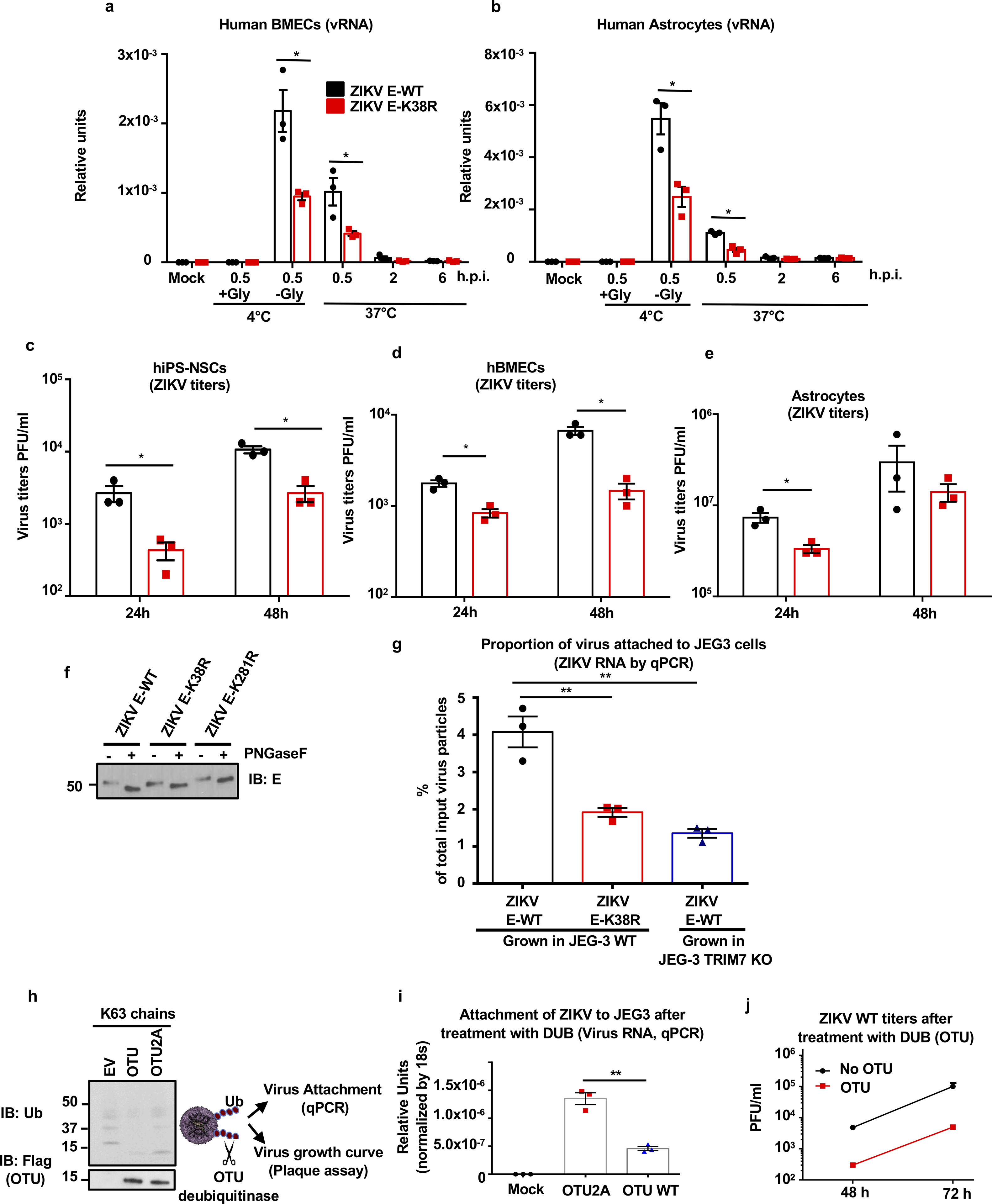
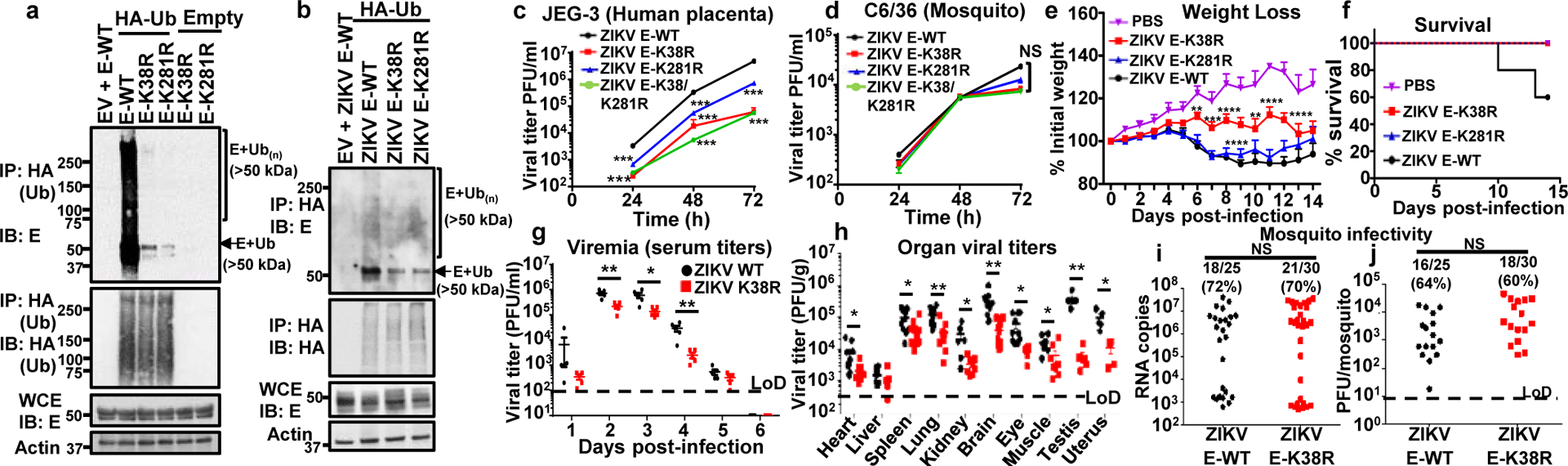
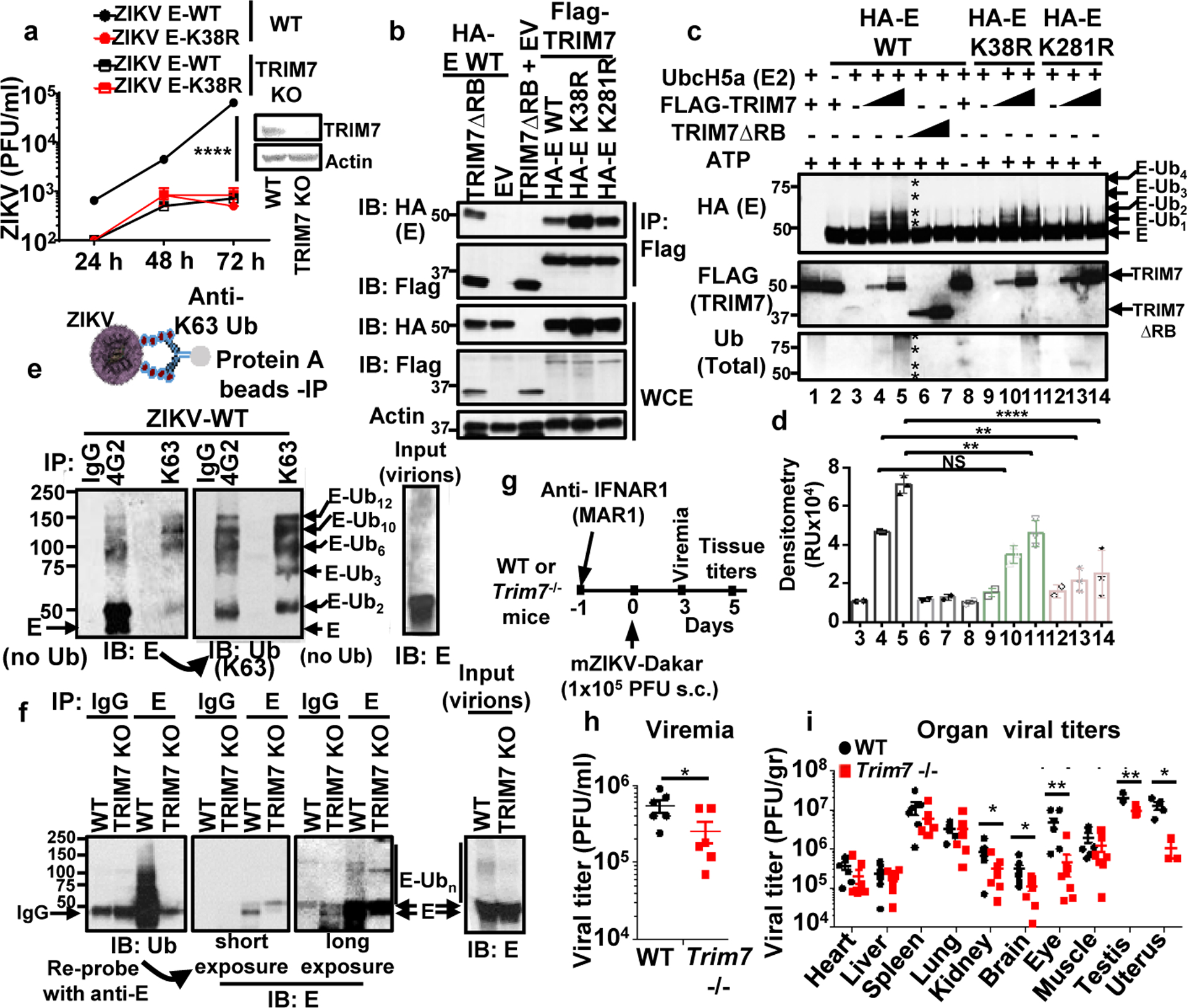
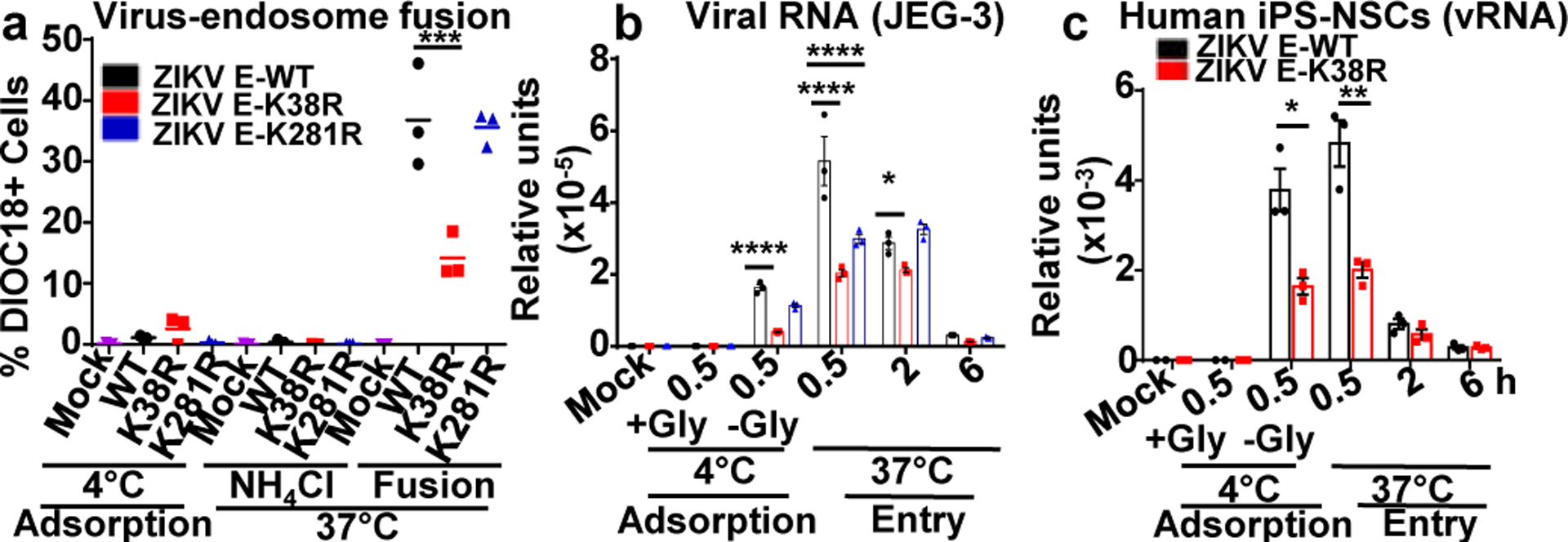
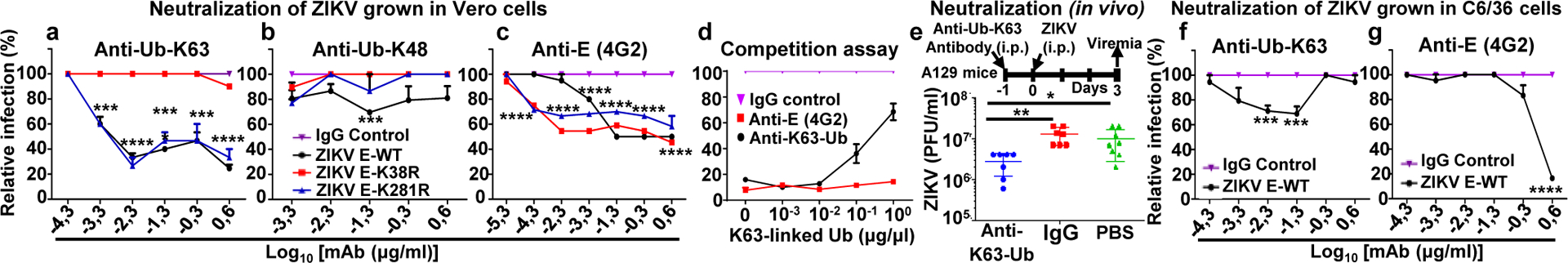
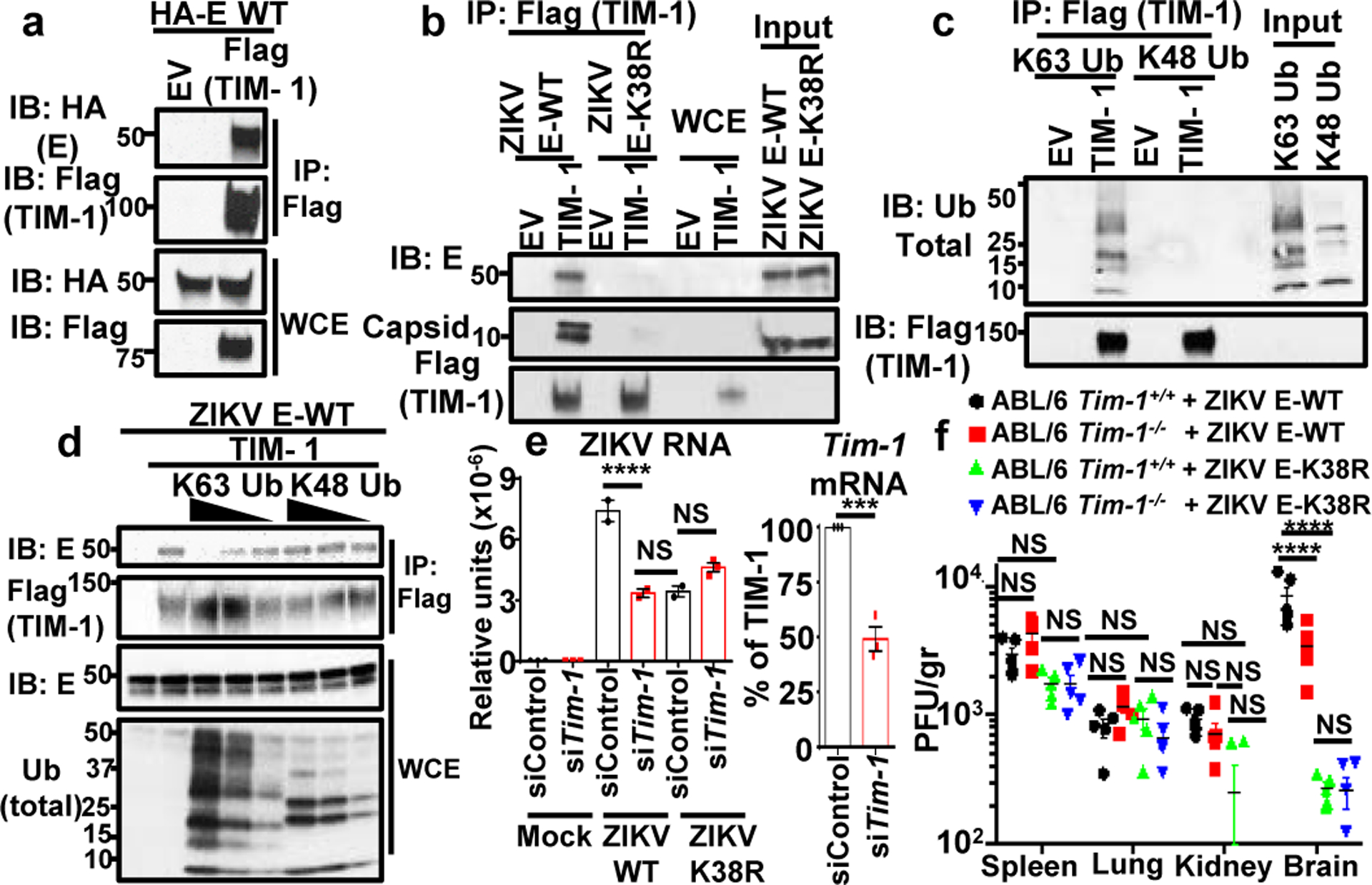
Zika virus (ZIKV) belongs to the family Flaviviridae, and is related to other viruses that cause human diseases. Unlike other flaviviruses, ZIKV infection can cause congenital neurological disorders and replicates efficiently in reproductive tissues1-3. Here we show that the envelope protein (E) of ZIKV is polyubiquitinated by the E3 ubiquitin ligase TRIM7 through Lys63 (K63)-linked polyubiquitination. Accordingly, ZIKV replicates less efficiently in the brain and reproductive tissues of Trim7-/- mice. Ubiquitinated E is present on infectious virions of ZIKV when they are released from specific cell types, and enhances virus attachment and entry into cells. Specifically, K63-linked polyubiquitin chains directly interact with the TIM1 (also known as HAVCR1) receptor of host cells, which enhances virus entry in cells as well as in brain tissue in vivo. Recombinant ZIKV mutants that lack ubiquitination are attenuated in human cells and in wild-type mice, but not in live mosquitoes. Monoclonal antibodies against K63-linked polyubiquitin specifically neutralize ZIKV and reduce viraemia in mice. Our results demonstrate that the ubiquitination of ZIKV E is an important determinant of virus entry, tropism and pathogenesis.
Conflict of interest statement
Figures
References
-
- Hills Susan L., Morgan Hennessey KR, Williams Charnetta, Oster Alexandra M., Fischer Marc, Mead Paul. Transmission of Zika Virus Through Sexual Contact with Travelers to Areas of Ongoing Transmission — Continental United States, 2016. CDC MMWR Morb Mortal Wkly Rep 65, 215–216 (2016). - PubMed
-
- Driggers RW et al. Zika Virus Infection with Prolonged Maternal Viremia and Fetal Brain Abnormalities. N Engl J Med 374, 2142–2151, doi:10.1056/NEJMoa1601824 (2016). - PubMed
-
- van Tol S, Hage A, Giraldo MI, Bharaj P & Rajsbaum R The TRIMendous Role of TRIMs in Virus-Host Interactions. Vaccines (Basel) 5, doi:10.3390/vaccines5030023 (2017). - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
