

Molecular profiles and immunomodulatory activities of glioblastoma-derived exosomes
- PMID: 32642708
- PMCID: PMC7262743
- DOI: 10.1093/noajnl/vdaa056
Molecular profiles and immunomodulatory activities of glioblastoma-derived exosomes
Abstract
Background: Glioblastoma is one of the most immunosuppressive human tumors. Emerging data suggest that glioblastoma-derived exosomes (GBex) reprogram the tumor microenvironment into a tumor-promoting milieu by mechanisms that not yet understood.
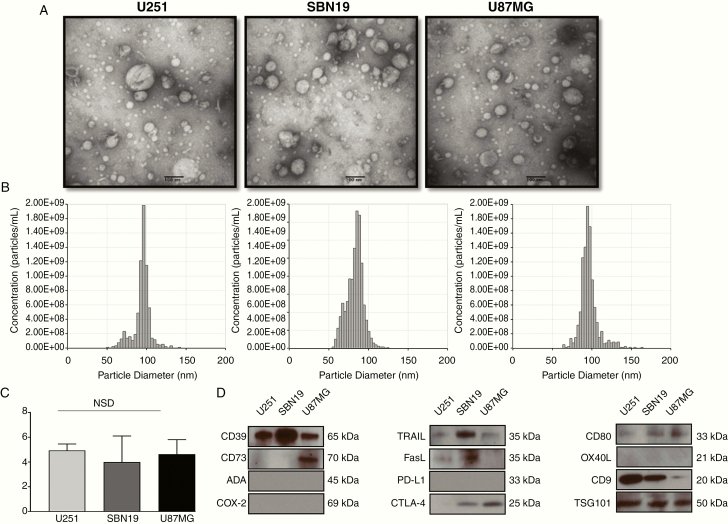
Methods: Exosomes were isolated from supernatants of glioblastoma cell lines by size exclusion chromatography. The GBex endosomal origin, size, protein cargos, and ex vivo effects on immune cell functions were determined. GBex were injected intravenously into mice to evaluate their ability to in vivo modulate normal immune cell subsets.
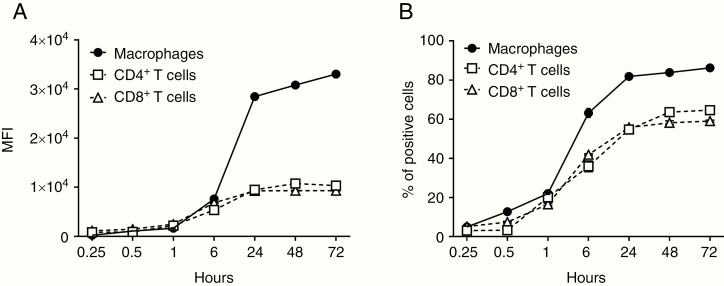
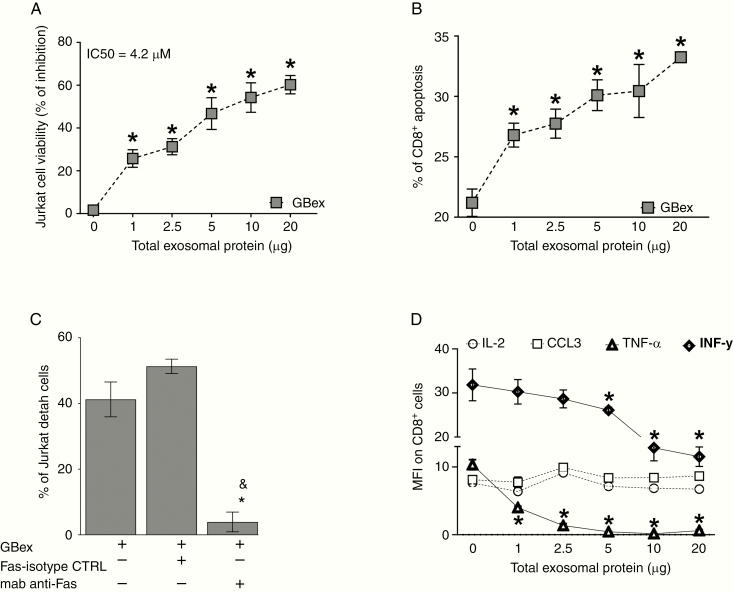
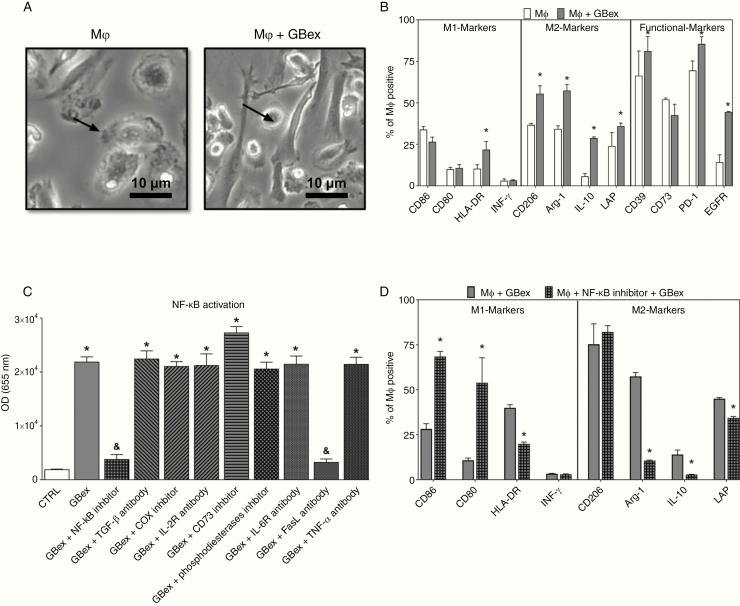
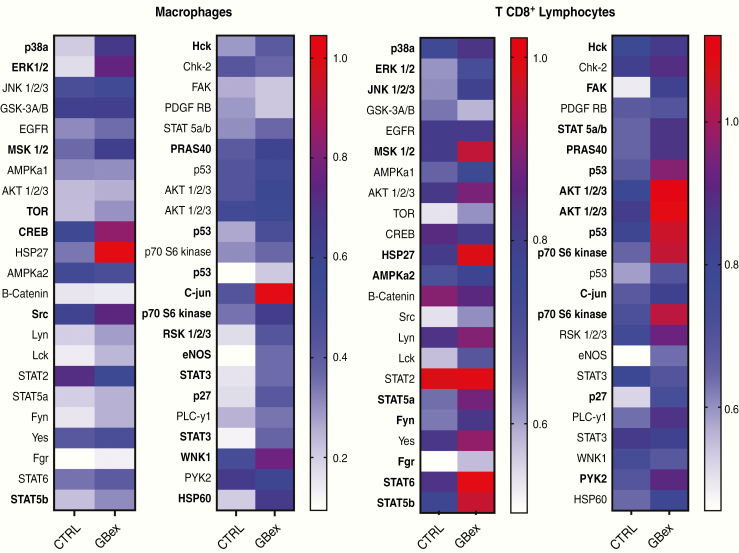
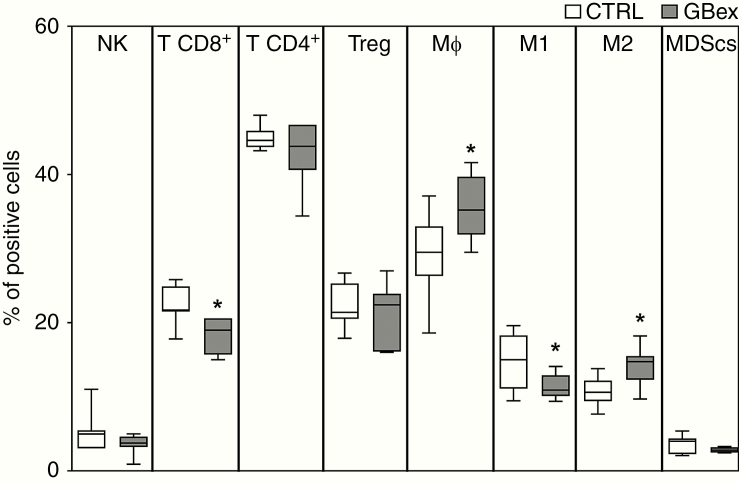
Results: GBex carried immunosuppressive proteins, including FasL, TRAIL, CTLA-4, CD39, and CD73, but contained few immunostimulatory proteins. GBex co-incubated with primary human immune cells induced simultaneous activation of multiple molecular pathways. In CD8+ T cells, GBex suppressed TNF-α and INF-γ release and mediated apoptosis. GBex suppressed natural killer (NK) and CD4+ T-cell activation. GBex activated the NF-κB pathway in macrophages and promoted their differentiation into M2 cells. Inhibition of the NF-κB pathway in macrophages reversed the GBex-mediated effects. GBex-driven reprogramming of macrophages involved the release of soluble factors that promoted tumor proliferation in vitro. In mice injected with GBex, the frequency of splenic CD8+ T cells, NK cells, and M1-like macrophages was reduced, while that of naïve and M2-like macrophages increased (P < .05).
Conclusions: GBex reprogrammed functions of all types of immune cells in vitro and altered their frequency in vivo. By creating and sustaining a highly immunosuppressive environment, GBex play a key role in promoting tumor progression.
Keywords: tumor microenvironment; exosomes; glioblastoma; immune system; macrophages.
© The Author(s) 2020. Published by Oxford University Press, the Society for Neuro-Oncology and the European Association of Neuro-Oncology.
Figures
References
-
- Stupp R, van den Bent MJ, Hegi ME. Optimal role of temozolomide in the treatment of malignant gliomas. Curr Neurol Neurosci Rep. 2005;5(3):198–206. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials