

Water-Gated Proton Transfer Dynamics in Respiratory Complex I
- PMID: 32643371
- PMCID: PMC7659035
- DOI: 10.1021/jacs.0c02789
Water-Gated Proton Transfer Dynamics in Respiratory Complex I
Abstract
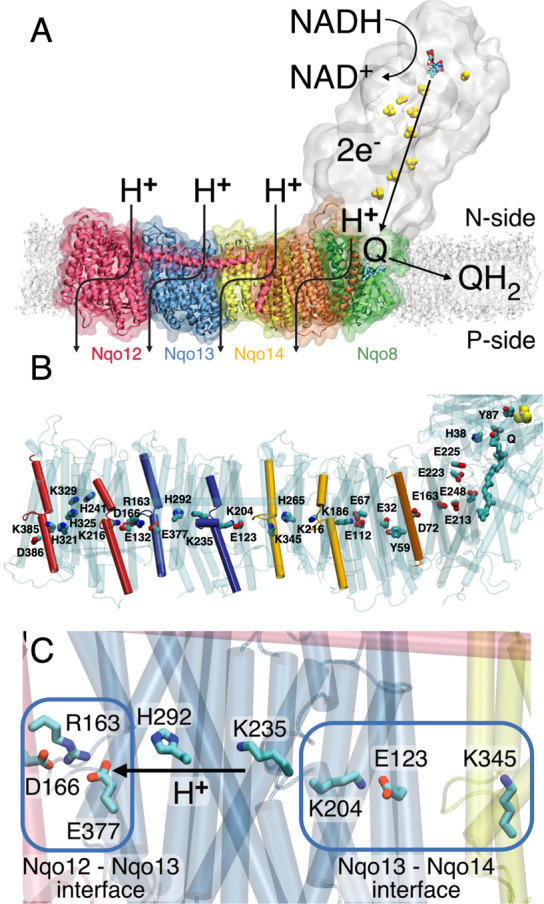
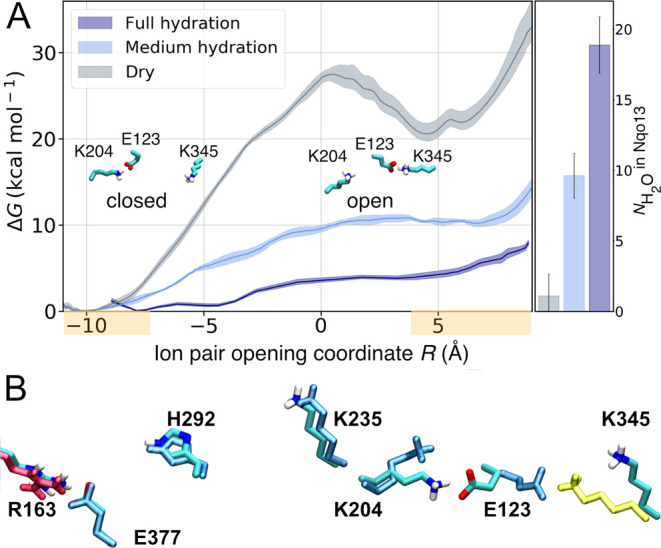
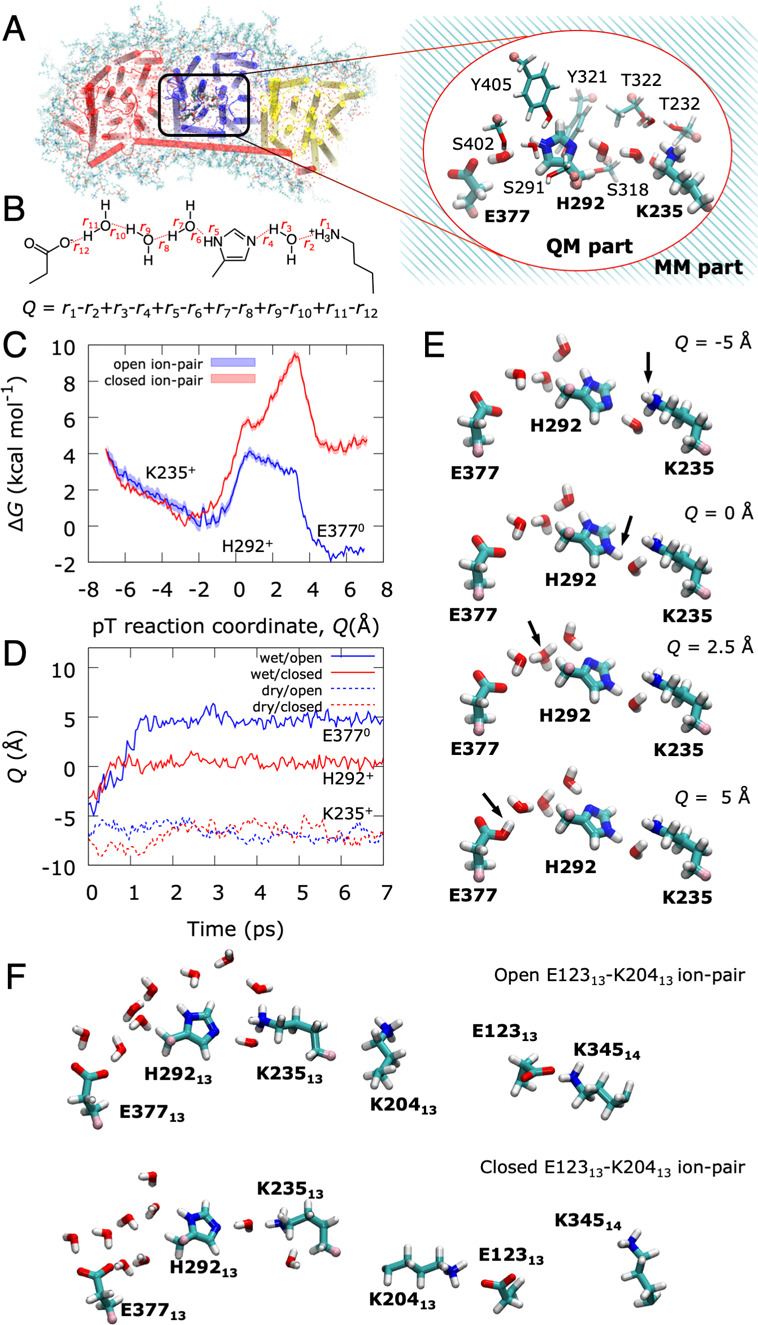
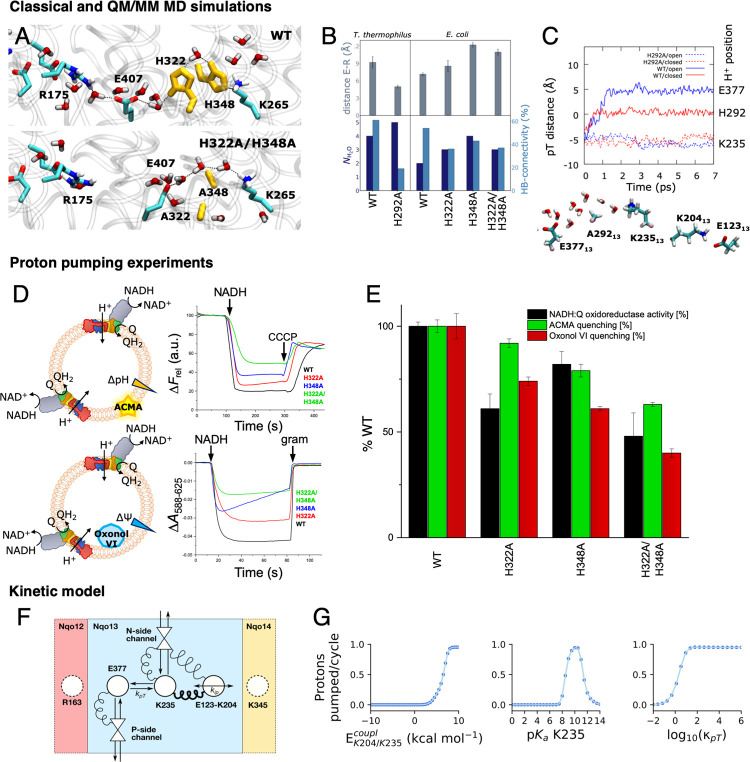
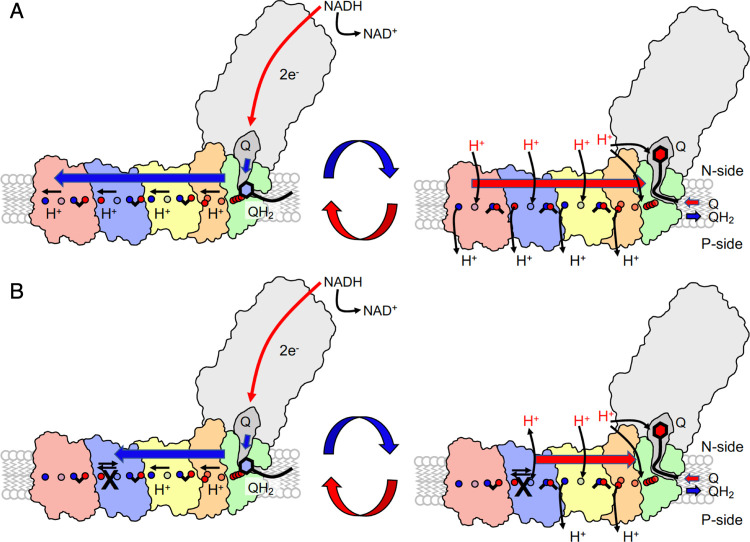
The respiratory complex I transduces redox energy into an electrochemical proton gradient in aerobic respiratory chains, powering energy-requiring processes in the cell. However, despite recently resolved molecular structures, the mechanism of this gigantic enzyme remains poorly understood. By combining large-scale quantum and classical simulations with site-directed mutagenesis and biophysical experiments, we show here how the conformational state of buried ion-pairs and water molecules control the protonation dynamics in the membrane domain of complex I and establish evolutionary conserved long-range coupling elements. We suggest that an electrostatic wave propagates in forward and reverse directions across the 200 Å long membrane domain during enzyme turnover, without significant dissipation of energy. Our findings demonstrate molecular principles that enable efficient long-range proton-electron coupling (PCET) and how perturbation of this PCET machinery may lead to development of mitochondrial disease.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
