

Implications of SARS-CoV-2 Mutations for Genomic RNA Structure and Host microRNA Targeting
- PMID: 32645951
- PMCID: PMC7370282
- DOI: 10.3390/ijms21134807
Implications of SARS-CoV-2 Mutations for Genomic RNA Structure and Host microRNA Targeting
Abstract
The SARS-CoV-2 virus is a recently-emerged zoonotic pathogen already well adapted to transmission and replication in humans. Although the mutation rate is limited, recently introduced mutations in SARS-CoV-2 have the potential to alter viral fitness. In addition to amino acid changes, mutations could affect RNA secondary structure critical to viral life cycle, or interfere with sequences targeted by host miRNAs. We have analysed subsets of genomes from SARS-CoV-2 isolates from around the globe and show that several mutations introduce changes in Watson-Crick pairing, with resultant changes in predicted secondary structure. Filtering to targets matching miRNAs expressed in SARS-CoV-2-permissive host cells, we identified ten separate target sequences in the SARS-CoV-2 genome; three of these targets have been lost through conserved mutations. A genomic site targeted by the highly abundant miR-197-5p, overexpressed in patients with cardiovascular disease, is lost by a conserved mutation. Our results are compatible with a model that SARS-CoV-2 replication within the human host is constrained by host miRNA defences. The impact of these and further mutations on secondary structures, miRNA targets or potential splice sites offers a new context in which to view future SARS-CoV-2 evolution, and a potential platform for engineering conditional attenuation to vaccine development, as well as providing a better understanding of viral tropism and pathogenesis.
Keywords: RNA secondary structure; SARS-CoV-2; conserved mutation; miRNA.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

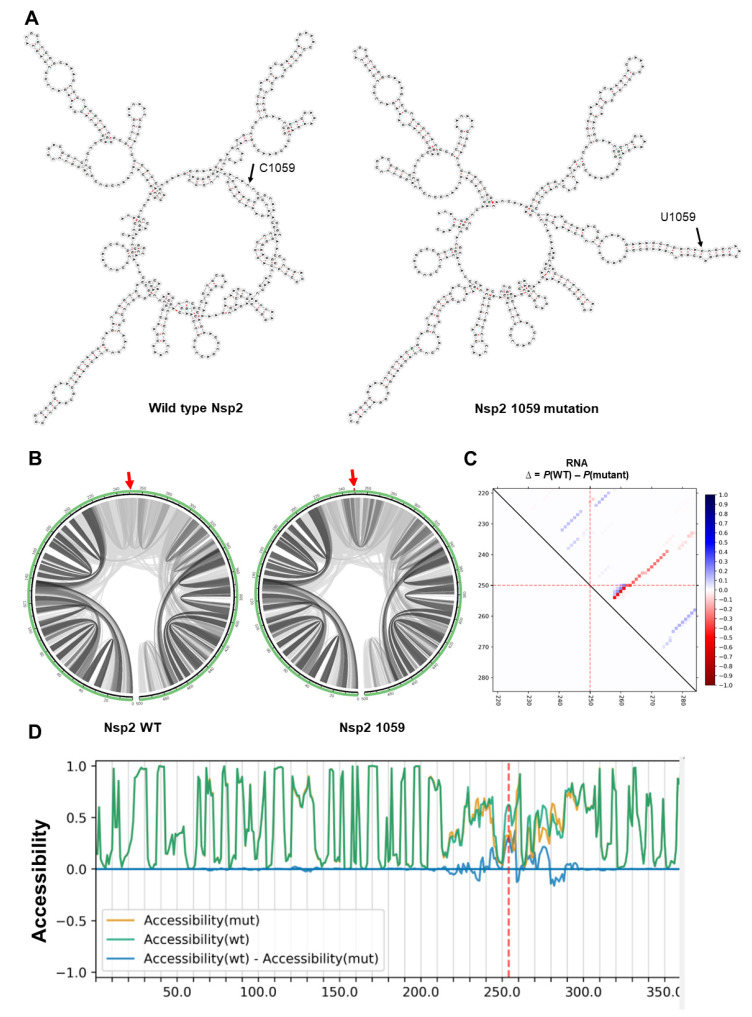
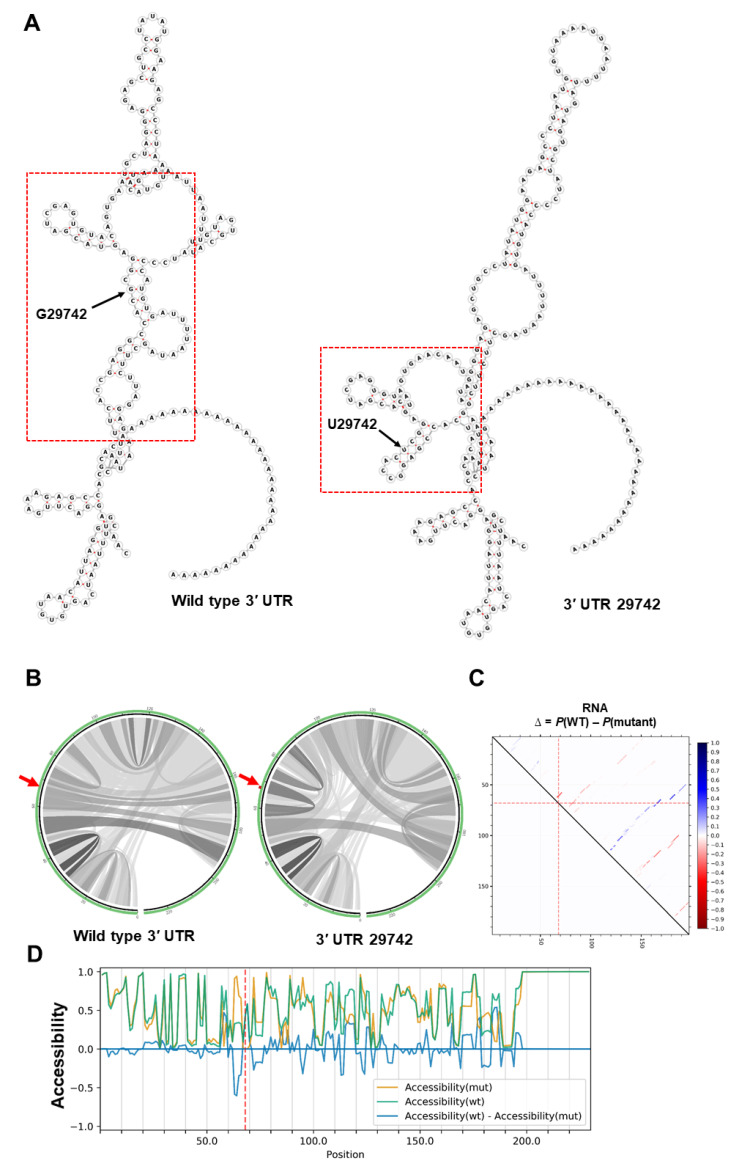
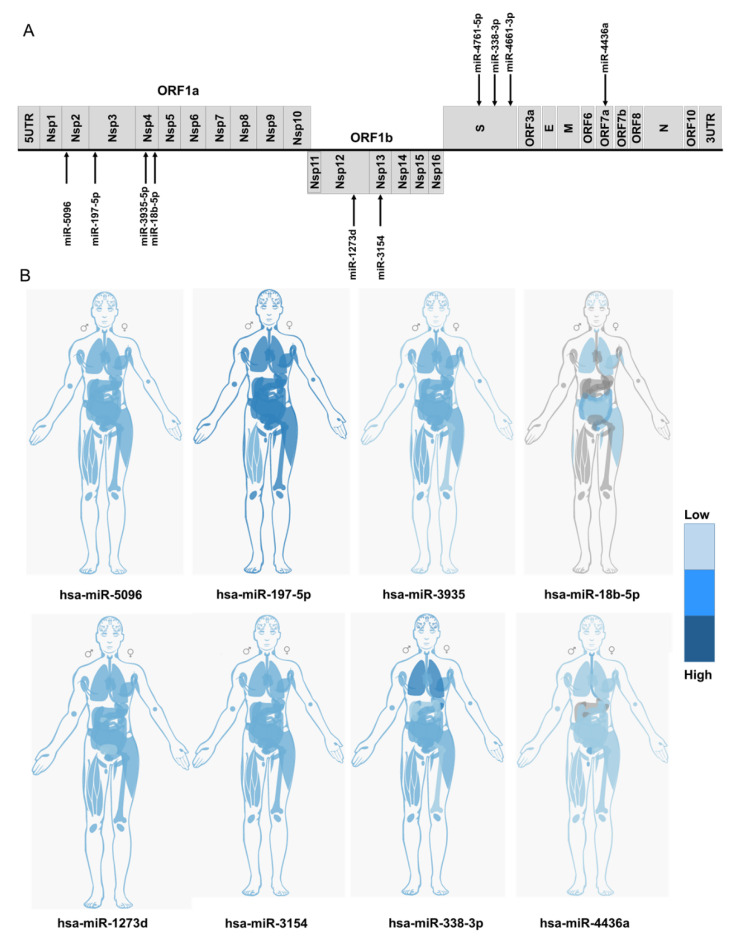
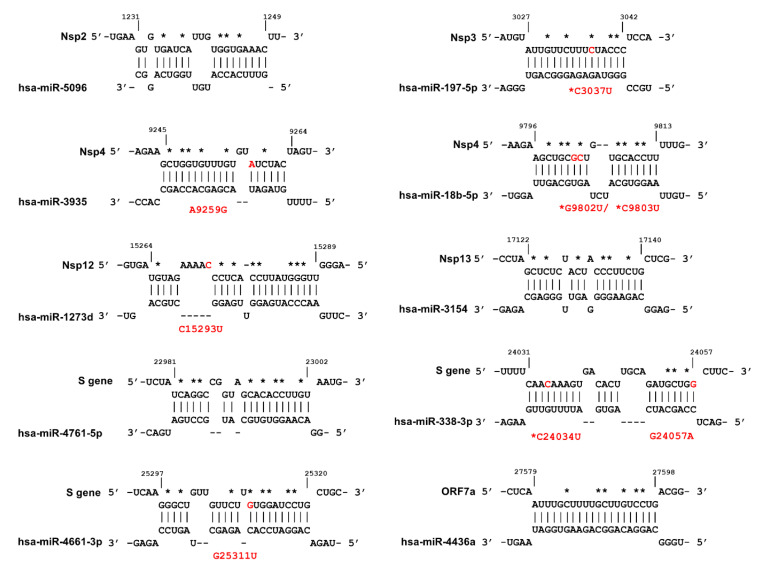

References
-
- Zhang L., Jackson C.B., Mou H., Ojha A., Rangarajan E.S., Izard T., Farzan M., Choe H. The D614G mutation in the SARS-CoV-2 spike protein reduces S1 shedding and increases infectivity. bioRxiv. 2020 doi: 10.1101/2020.06.12.148726. - DOI
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
