

Lysines in the lyase active site of DNA polymerase β destabilize nonspecific DNA binding, facilitating searching and DNA gap recognition
- PMID: 32647014
- PMCID: PMC7443498
- DOI: 10.1074/jbc.RA120.013547
Lysines in the lyase active site of DNA polymerase β destabilize nonspecific DNA binding, facilitating searching and DNA gap recognition
Abstract
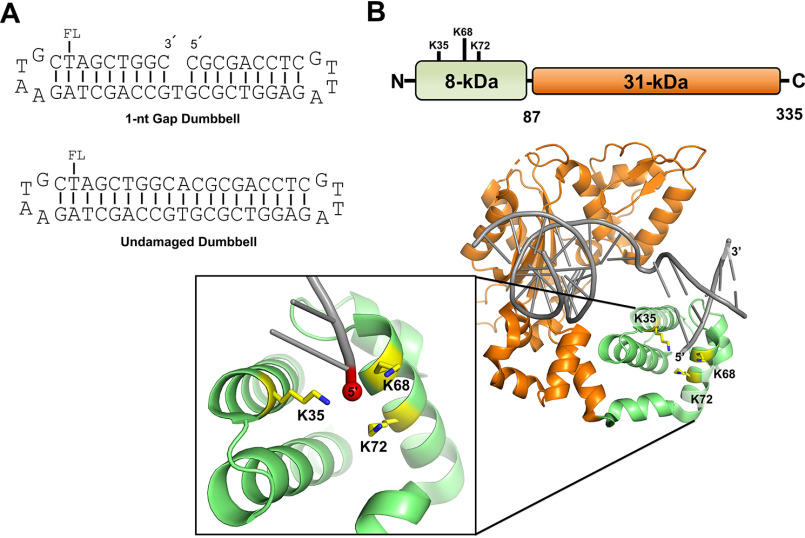
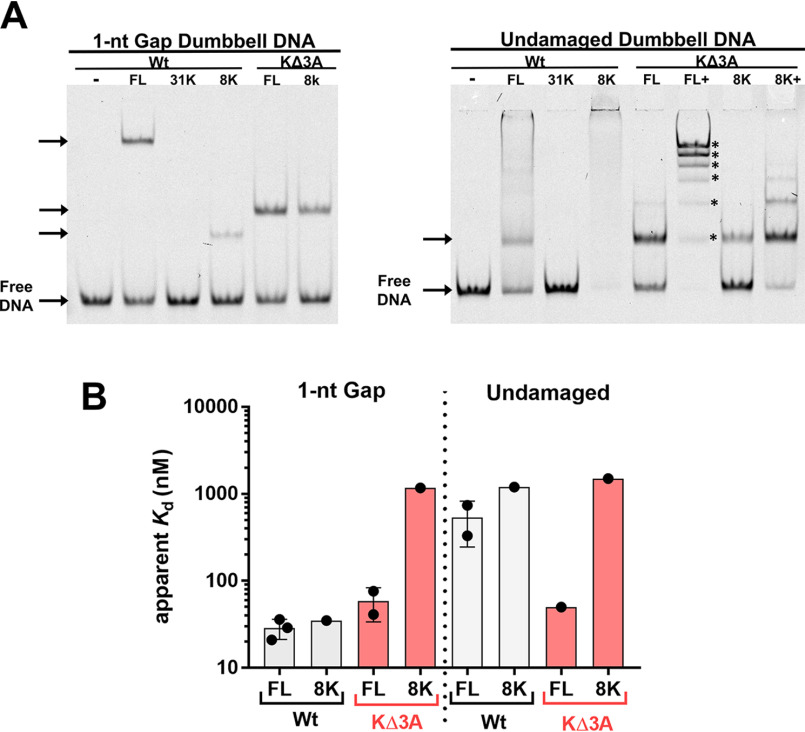
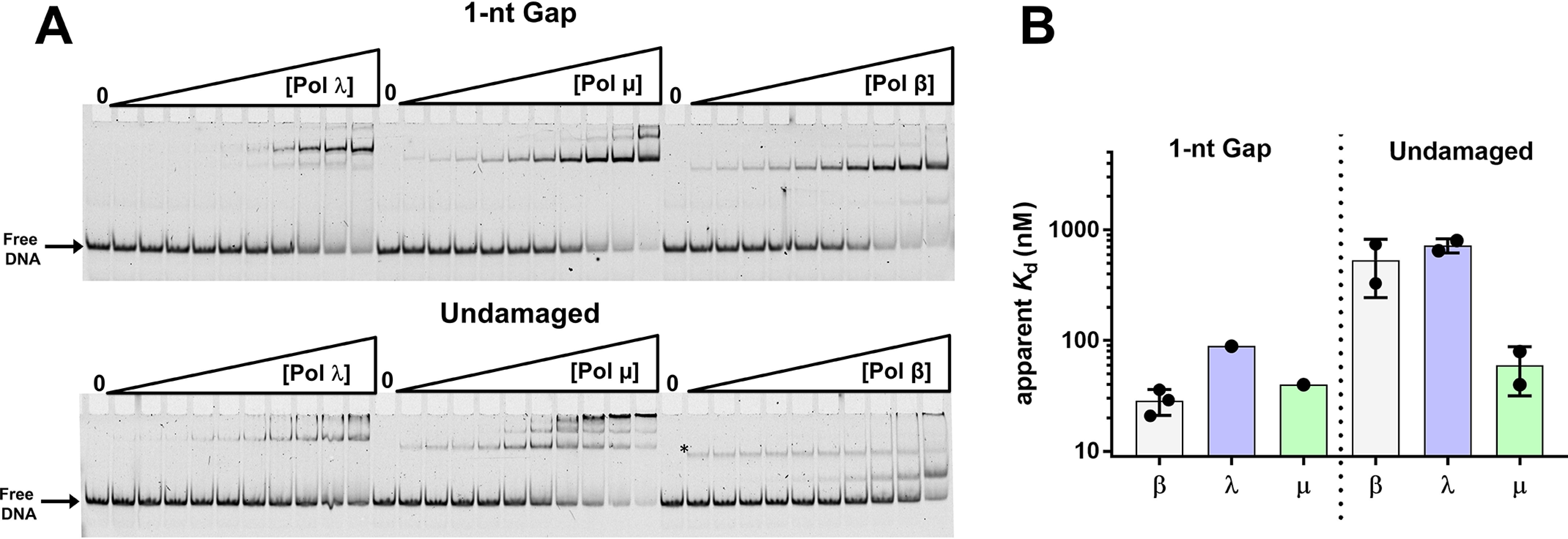
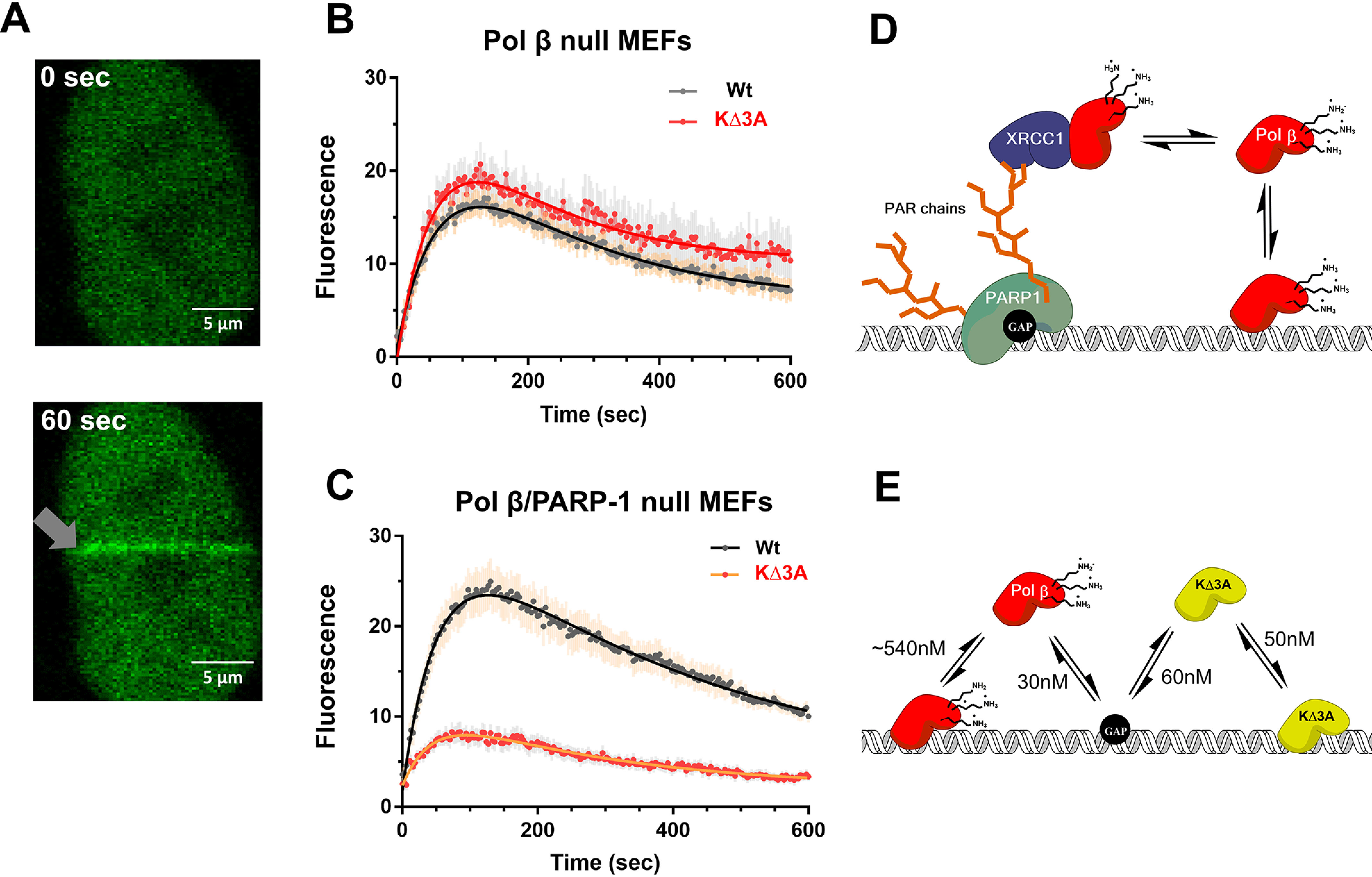
DNA polymerase (pol) β catalyzes two reactions at DNA gaps generated during base excision repair, gap-filling DNA synthesis and lyase-dependent 5´-end deoxyribose phosphate removal. The lyase domain of pol β has been proposed to function in DNA gap recognition and to facilitate DNA scanning during substrate search. However, the mechanisms and molecular interactions used by pol β for substrate search and recognition are not clear. To provide insight into this process, a comparison was made of the DNA binding affinities of WT pol β, pol λ, and pol μ, and several variants of pol β, for 1-nt-gap-containing and undamaged DNA. Surprisingly, this analysis revealed that mutation of three lysine residues in the lyase active site of pol β, 35, 68, and 72, to alanine (pol β KΔ3A) increased the binding affinity for nonspecific DNA ∼11-fold compared with that of the WT. WT pol μ, lacking homologous lysines, displayed nonspecific DNA binding behavior similar to that of pol β KΔ3A, in line with previous data demonstrating both enzymes were deficient in processive searching. In fluorescent microscopy experiments using mouse fibroblasts deficient in PARP-1, the ability of pol β KΔ3A to localize to sites of laser-induced DNA damage was strongly decreased compared with that of WT pol β. These data suggest that the three lysines in the lyase active site destabilize pol β when bound to DNA nonspecifically, promoting DNA scanning and providing binding specificity for gapped DNA.
Keywords: DNA binding protein; DNA binding proteins; DNA damage; DNA polymerase; DNA repair; DNA–protein interaction; base excision repair (BER); facilitated diffusion; nonspecific DNA binding; processive search.
Conflict of interest statement
Conflict of interest—The authors declare that they have no conflicts of interest with the contents of this article.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
