

A Novel GATA2 Protein Reporter Mouse Reveals Hematopoietic Progenitor Cell Types
- PMID: 32649900
- PMCID: PMC7419669
- DOI: 10.1016/j.stemcr.2020.06.008
A Novel GATA2 Protein Reporter Mouse Reveals Hematopoietic Progenitor Cell Types
Abstract
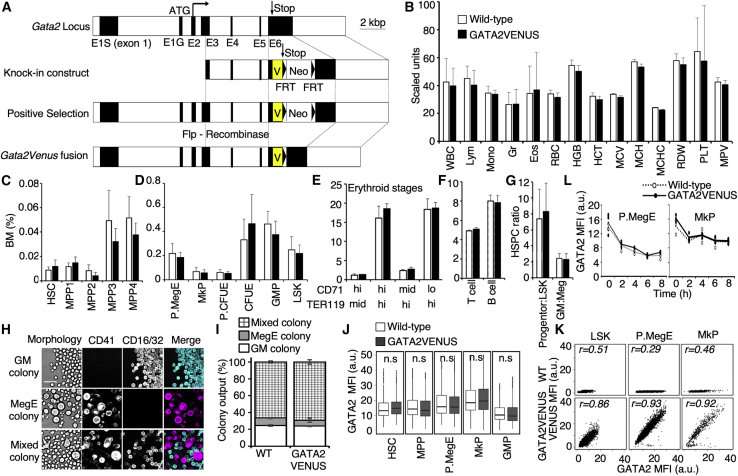
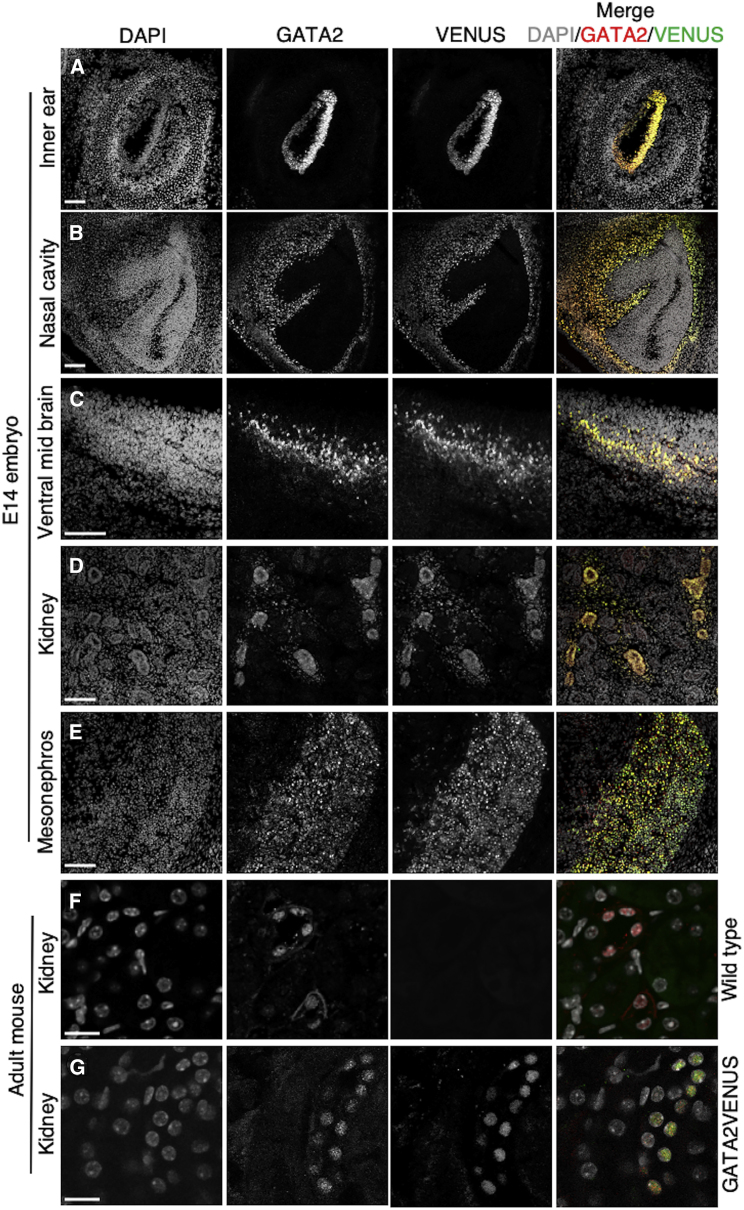
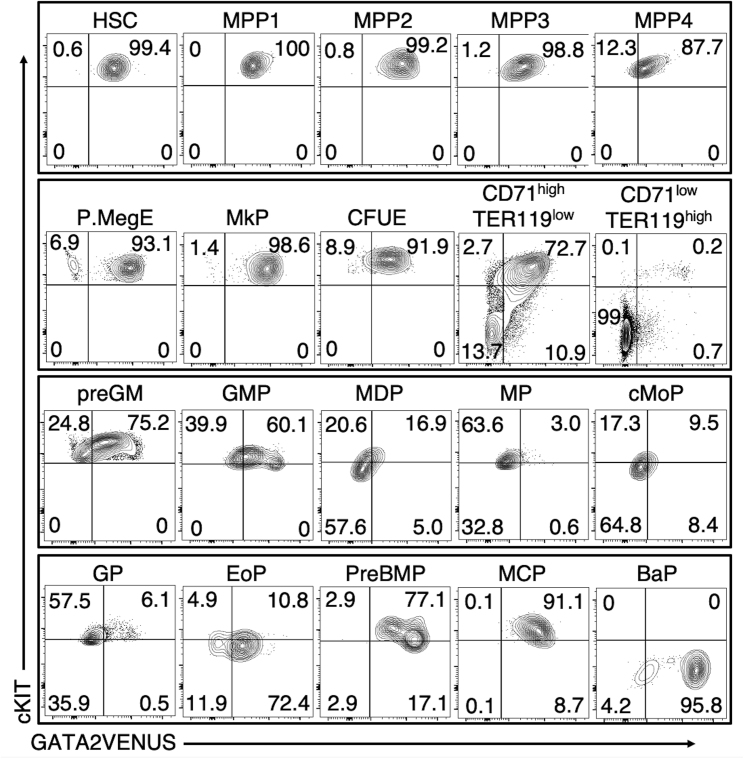
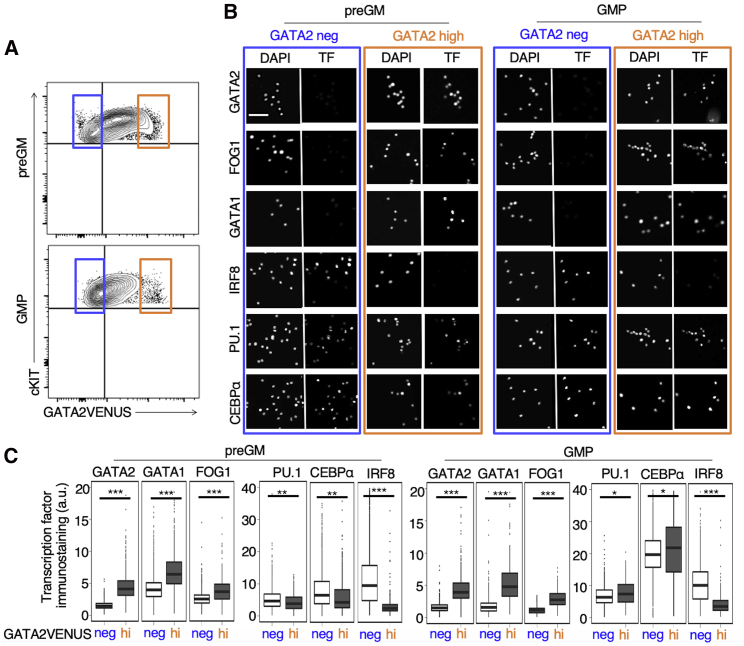
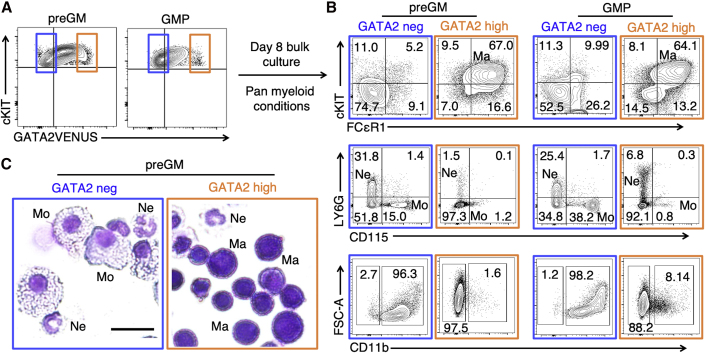
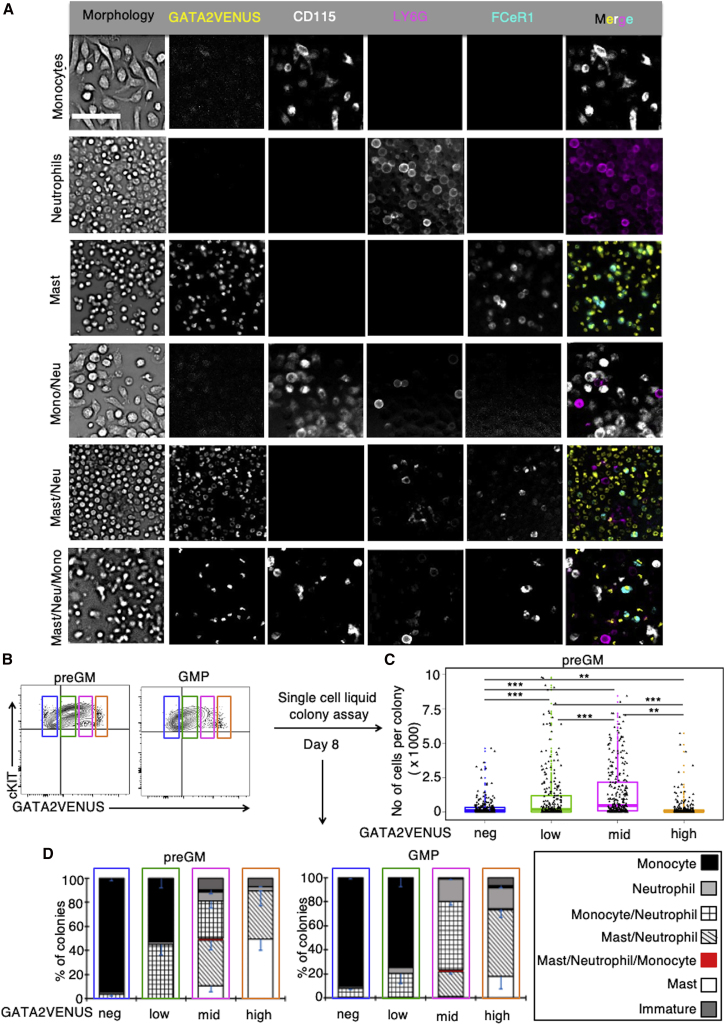
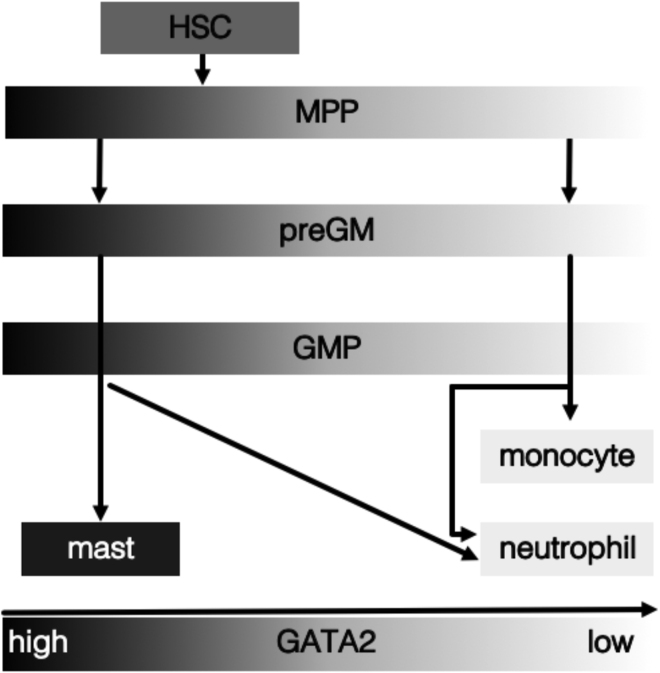
The transcription factor (TF) GATA2 plays a key role in organ development and cell fate control in the central nervous, urogenital, respiratory, and reproductive systems, and in primitive and definitive hematopoiesis. Here, we generate a knockin protein reporter mouse line expressing a GATA2VENUS fusion from the endogenous Gata2 genomic locus, with correct expression and localization of GATA2VENUS in different organs. GATA2VENUS expression is heterogeneous in different hematopoietic stem and progenitor cell populations (HSPCs), identifies functionally distinct subsets, and suggests a novel monocyte and mast cell lineage bifurcation point. GATA2 levels further correlate with proliferation and lineage outcome of hematopoietic progenitors. The GATA2VENUS mouse line improves the identification of specific live cell types during embryonic and adult development and will be crucial for analyzing GATA2 protein dynamics in TF networks.
Keywords: GATA2; cell fate; development; fluorescent protein; hematopoietic stem and progenitor cell; monocyte and mast cell lineage; transcription factor networks; transgenic mouse.
Copyright © 2020 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Akashi K., Traver D., Miyamoto T., Weissman I.L. A clonogenic common myeloid progenitor that gives rise to all myeloid lineages. Nature. 2000;404:193–197. - PubMed
-
- Bresnick E.H., Martowicz M., Pal S., Johnson K.D. Developmental control via GATA factor interplay at chromatin domains. J. Cell. Physiol. 2005;205:1–9. - PubMed
-
- Cabezas-Wallscheid N., Klimmeck D., Hansson J., Lipka D.B., Reyes A., Wang Q., Weichenhan D., Lier A., Von Paleske L., Renders S. Identification of regulatory networks in HSCs and their immediate progeny via integrated proteome, transcriptome, and DNA methylome analysis. Cell Stem Cell. 2014;15:507–522. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
