

Chemosensory mechanisms of host seeking and infectivity in skin-penetrating nematodes
- PMID: 32651273
- PMCID: PMC7395504
- DOI: 10.1073/pnas.1909710117
Chemosensory mechanisms of host seeking and infectivity in skin-penetrating nematodes
Abstract
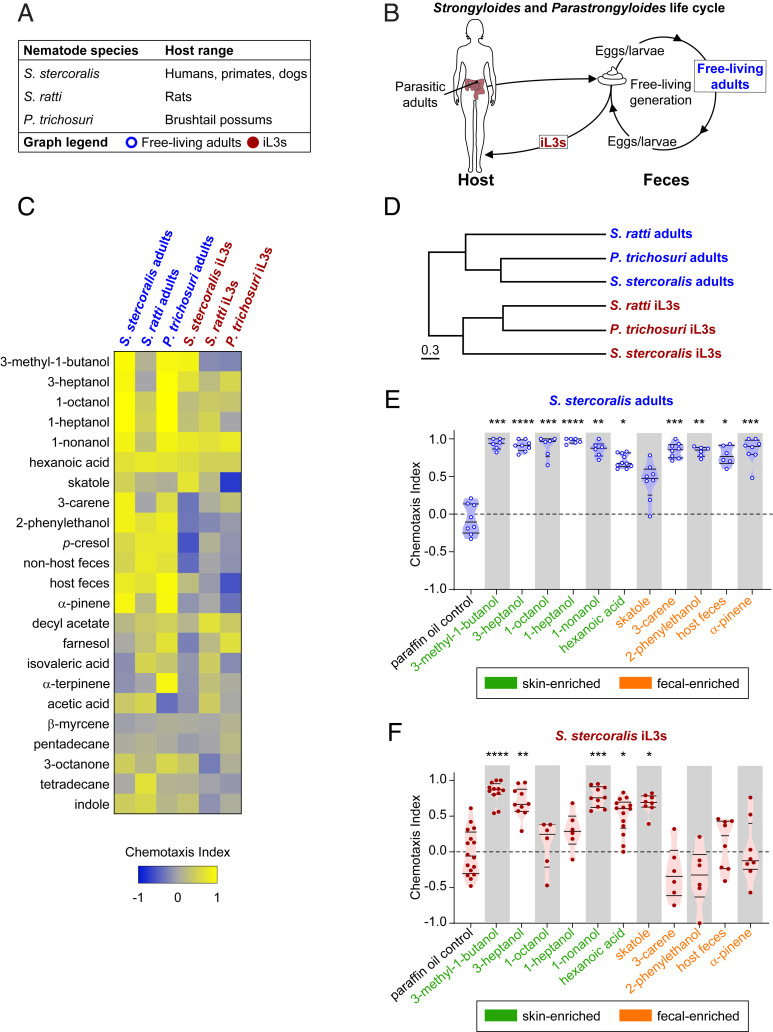
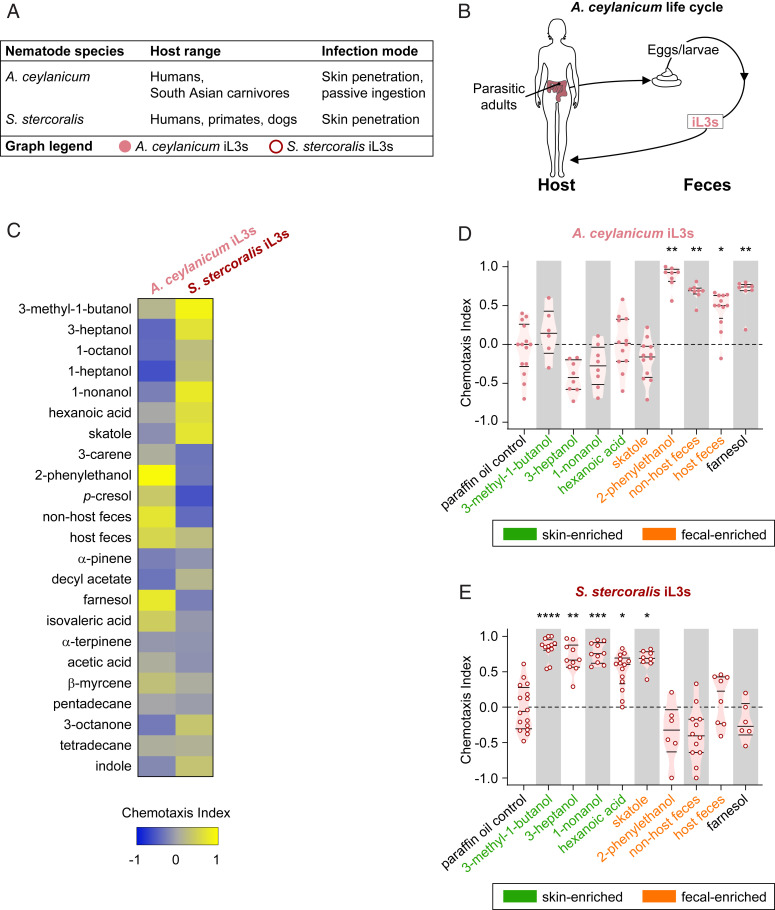
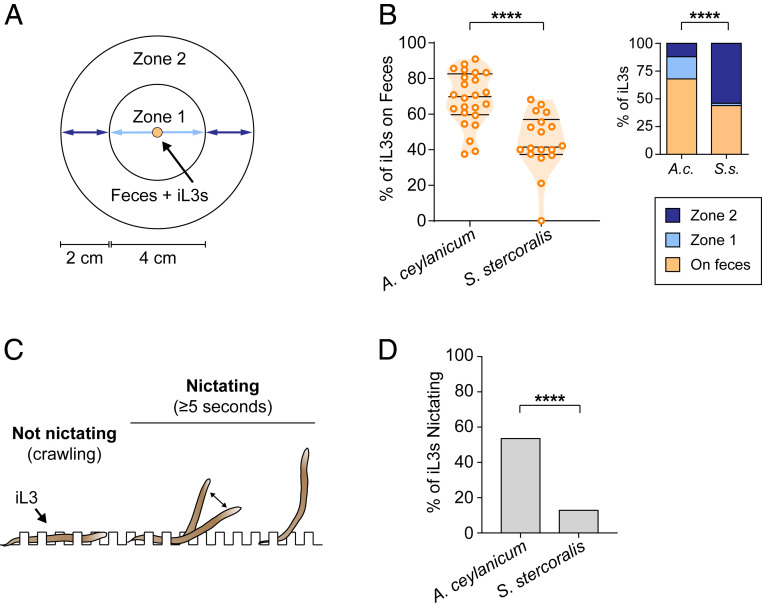
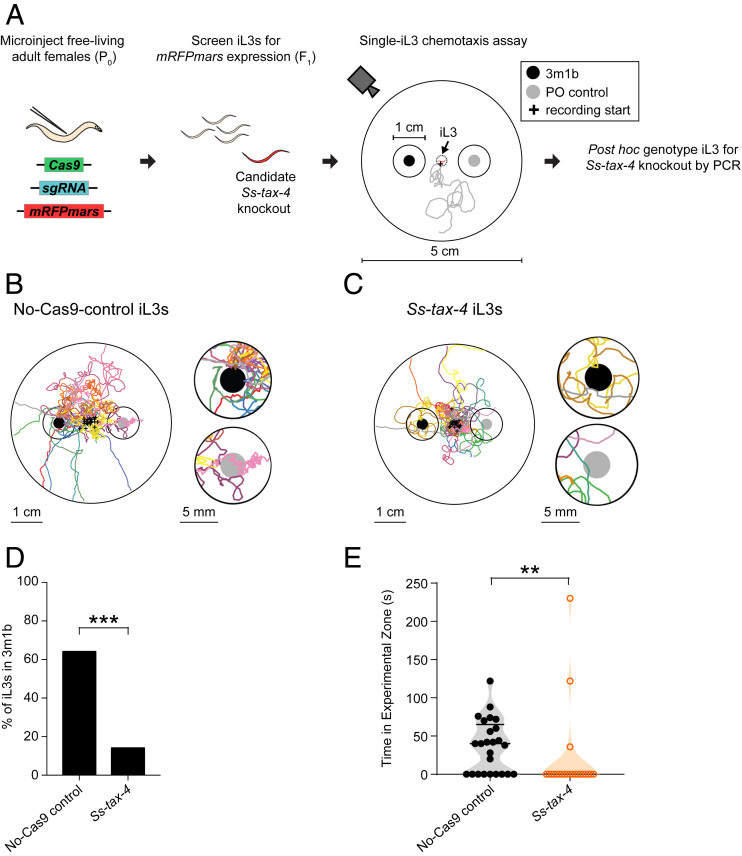
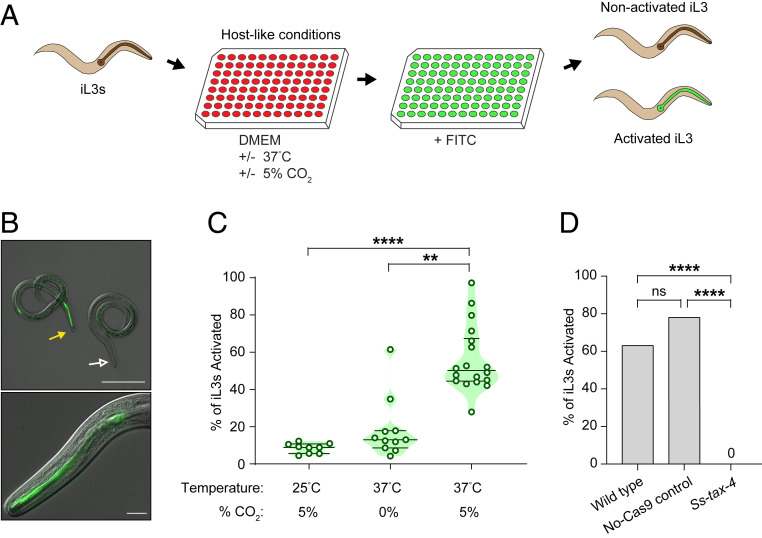
Approximately 800 million people worldwide are infected with one or more species of skin-penetrating nematodes. These parasites persist in the environment as developmentally arrested third-stage infective larvae (iL3s) that navigate toward host-emitted cues, contact host skin, and penetrate the skin. iL3s then reinitiate development inside the host in response to sensory cues, a process called activation. Here, we investigate how chemosensation drives host seeking and activation in skin-penetrating nematodes. We show that the olfactory preferences of iL3s are categorically different from those of free-living adults, which may restrict host seeking to iL3s. The human-parasitic threadworm Strongyloides stercoralis and hookworm Ancylostoma ceylanicum have highly dissimilar olfactory preferences, suggesting that these two species may use distinct strategies to target humans. CRISPR/Cas9-mediated mutagenesis of the S. stercoralis tax-4 gene abolishes iL3 attraction to a host-emitted odorant and prevents activation. Our results suggest an important role for chemosensation in iL3 host seeking and infectivity and provide insight into the molecular mechanisms that underlie these processes.
Keywords: Strongyloides stercoralis; chemosensation; host seeking; parasitic helminth; parasitic nematode.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Traub R. J., Ancylostoma ceylanicum, a re-emerging but neglected parasitic zoonosis. Int. J. Parasitol. 43, 1009–1015 (2013). - PubMed
