

Human GTPBP5 (MTG2) fuels mitoribosome large subunit maturation by facilitating 16S rRNA methylation
- PMID: 32652011
- PMCID: PMC7430652
- DOI: 10.1093/nar/gkaa592
Human GTPBP5 (MTG2) fuels mitoribosome large subunit maturation by facilitating 16S rRNA methylation
Abstract
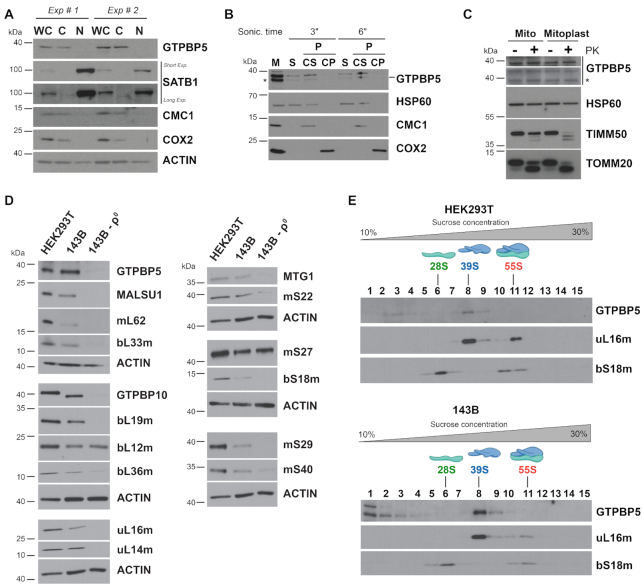
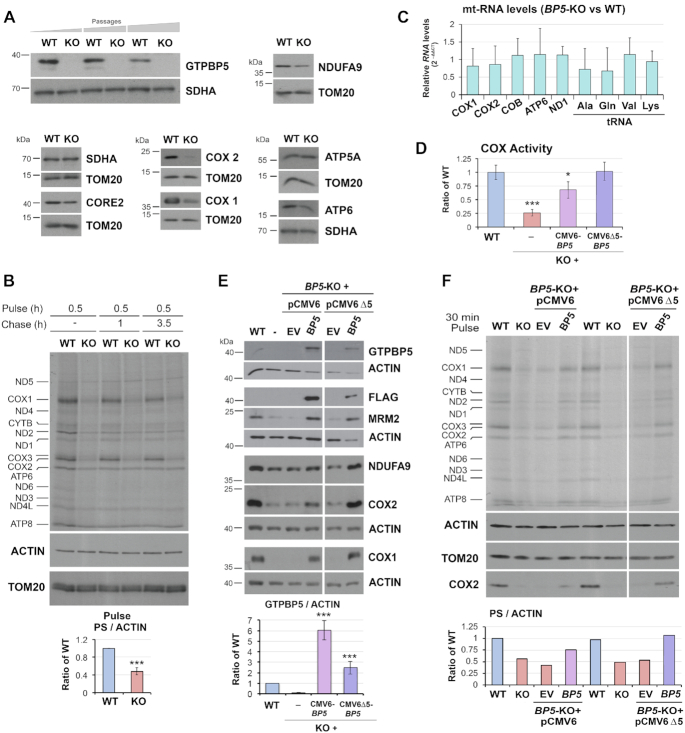
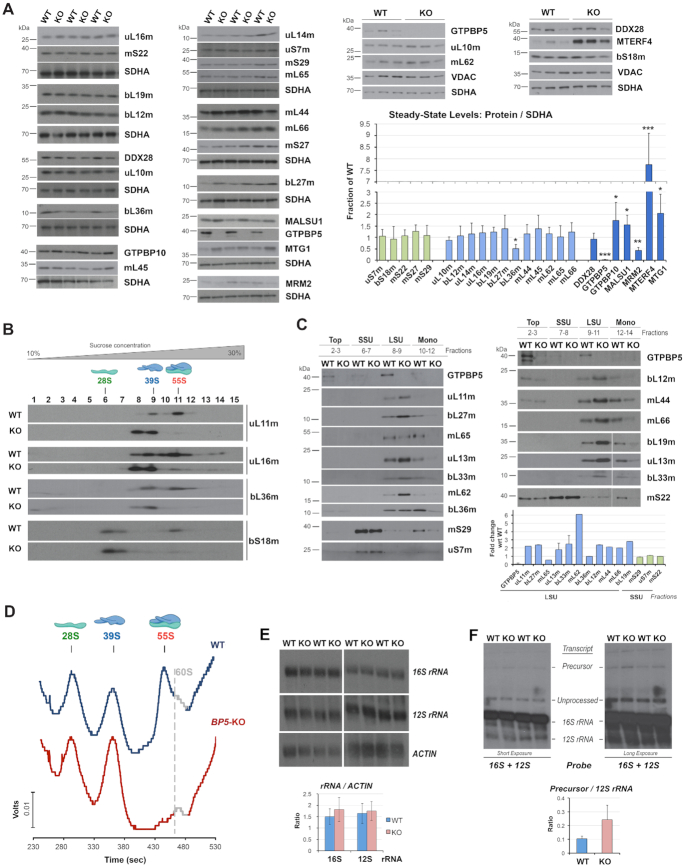
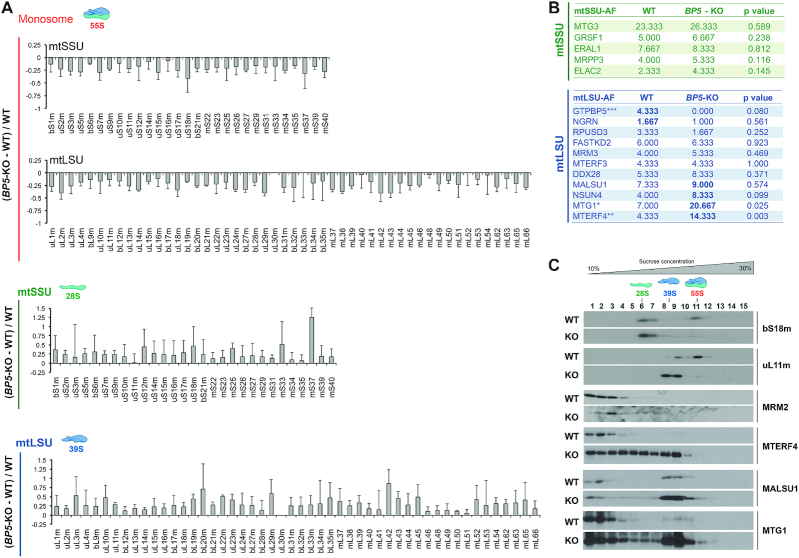
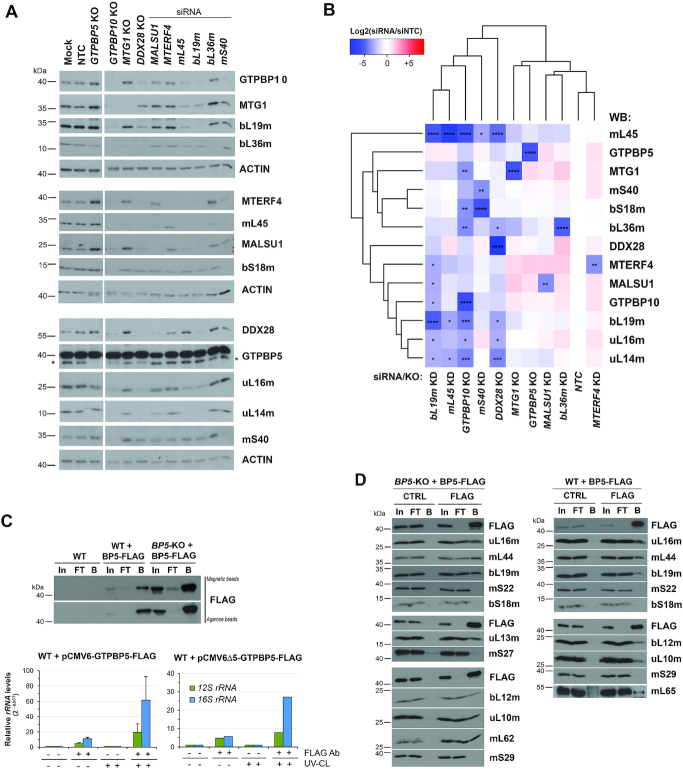
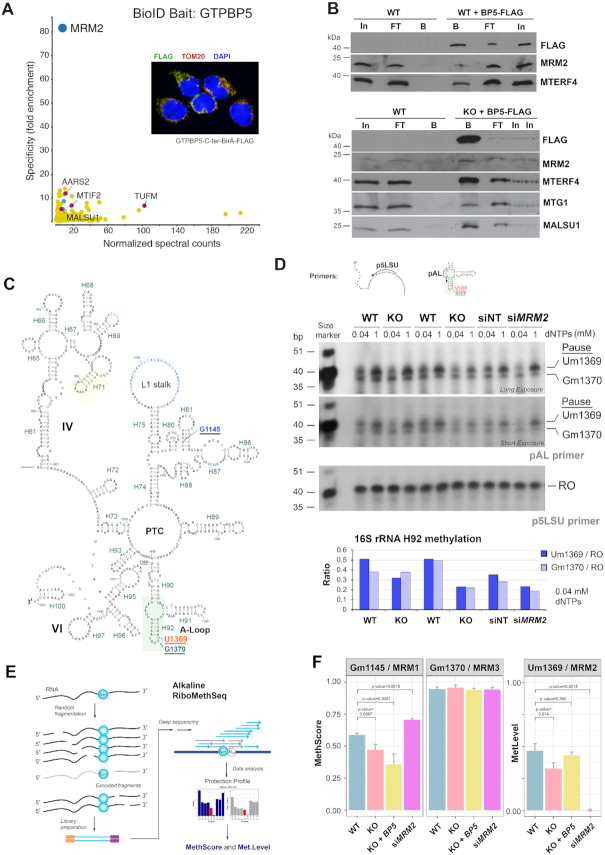
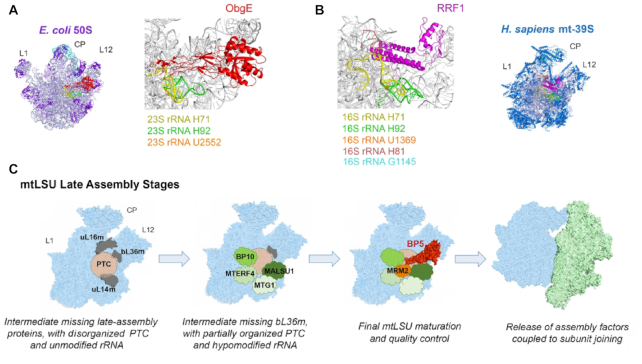
Biogenesis of mammalian mitochondrial ribosomes (mitoribosomes) involves several conserved small GTPases. Here, we report that the Obg family protein GTPBP5 or MTG2 is a mitochondrial protein whose absence in a TALEN-induced HEK293T knockout (KO) cell line leads to severely decreased levels of the 55S monosome and attenuated mitochondrial protein synthesis. We show that a fraction of GTPBP5 co-sediments with the large mitoribosome subunit (mtLSU), and crosslinks specifically with the 16S rRNA, and several mtLSU proteins and assembly factors. Notably, the latter group includes MTERF4, involved in monosome assembly, and MRM2, the methyltransferase that catalyzes the modification of the 16S mt-rRNA A-loop U1369 residue. The GTPBP5 interaction with MRM2 was also detected using the proximity-dependent biotinylation (BioID) assay. In GTPBP5-KO mitochondria, the mtLSU lacks bL36m, accumulates an excess of the assembly factors MTG1, GTPBP10, MALSU1 and MTERF4, and contains hypomethylated 16S rRNA. We propose that GTPBP5 primarily fuels proper mtLSU maturation by securing efficient methylation of two 16S rRNA residues, and ultimately serves to coordinate subunit joining through the release of late-stage mtLSU assembly factors. In this way, GTPBP5 provides an ultimate quality control checkpoint function during mtLSU assembly that minimizes premature subunit joining to ensure the assembly of the mature 55S monosome.
© The Author(s) 2020. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
Similar articles
-
Human GTPBP5 is involved in the late stage of mitoribosome large subunit assembly.Nucleic Acids Res. 2021 Jan 11;49(1):354-370. doi: 10.1093/nar/gkaa1131. Nucleic Acids Res. 2021. PMID: 33283228 Free PMC article.
-
Human GTPBP10 is required for mitoribosome maturation.Nucleic Acids Res. 2018 Nov 30;46(21):11423-11437. doi: 10.1093/nar/gky938. Nucleic Acids Res. 2018. PMID: 30321378 Free PMC article.
-
The human Obg protein GTPBP10 is involved in mitoribosomal biogenesis.Nucleic Acids Res. 2018 Sep 19;46(16):8471-8482. doi: 10.1093/nar/gky701. Nucleic Acids Res. 2018. PMID: 30085210 Free PMC article.
-
Role of GTPases in Driving Mitoribosome Assembly.Trends Cell Biol. 2021 Apr;31(4):284-297. doi: 10.1016/j.tcb.2020.12.008. Epub 2021 Jan 5. Trends Cell Biol. 2021. PMID: 33419649 Free PMC article. Review.
-
Insights into mitoribosomal biogenesis from recent structural studies.Trends Biochem Sci. 2023 Jul;48(7):629-641. doi: 10.1016/j.tibs.2023.04.002. Epub 2023 May 10. Trends Biochem Sci. 2023. PMID: 37169615 Review.
Cited by
-
Dual function of GTPBP6 in biogenesis and recycling of human mitochondrial ribosomes.Nucleic Acids Res. 2020 Dec 16;48(22):12929-12942. doi: 10.1093/nar/gkaa1132. Nucleic Acids Res. 2020. PMID: 33264405 Free PMC article.
-
WBSCR16 is essential for mitochondrial 16S rRNA processing in mammals.Nucleic Acids Res. 2025 Jan 24;53(3):gkae1325. doi: 10.1093/nar/gkae1325. Nucleic Acids Res. 2025. PMID: 39878214 Free PMC article.
-
Structural insights into the role of GTPBP10 in the RNA maturation of the mitoribosome.Nat Commun. 2023 Dec 2;14(1):7991. doi: 10.1038/s41467-023-43599-z. Nat Commun. 2023. PMID: 38042949 Free PMC article.
-
Structural basis for late maturation steps of the human mitoribosomal large subunit.Nat Commun. 2021 Jun 16;12(1):3673. doi: 10.1038/s41467-021-23617-8. Nat Commun. 2021. PMID: 34135318 Free PMC article.
-
Molecular pathways in mitochondrial disorders due to a defective mitochondrial protein synthesis.Front Cell Dev Biol. 2024 May 24;12:1410245. doi: 10.3389/fcell.2024.1410245. eCollection 2024. Front Cell Dev Biol. 2024. PMID: 38855161 Free PMC article. Review.
References
-
- Greber B.J., Bieri P., Leibundgut M., Leitner A., Aebersold R., Boehringer D., Ban N.. Ribosome. The complete structure of the 55S mammalian mitochondrial ribosome. Science. 2015; 348:303–308. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
