

The Cerebellar Nuclei and Dexterous Limb Movements
- PMID: 32652173
- PMCID: PMC7688491
- DOI: 10.1016/j.neuroscience.2020.06.046
The Cerebellar Nuclei and Dexterous Limb Movements
Abstract
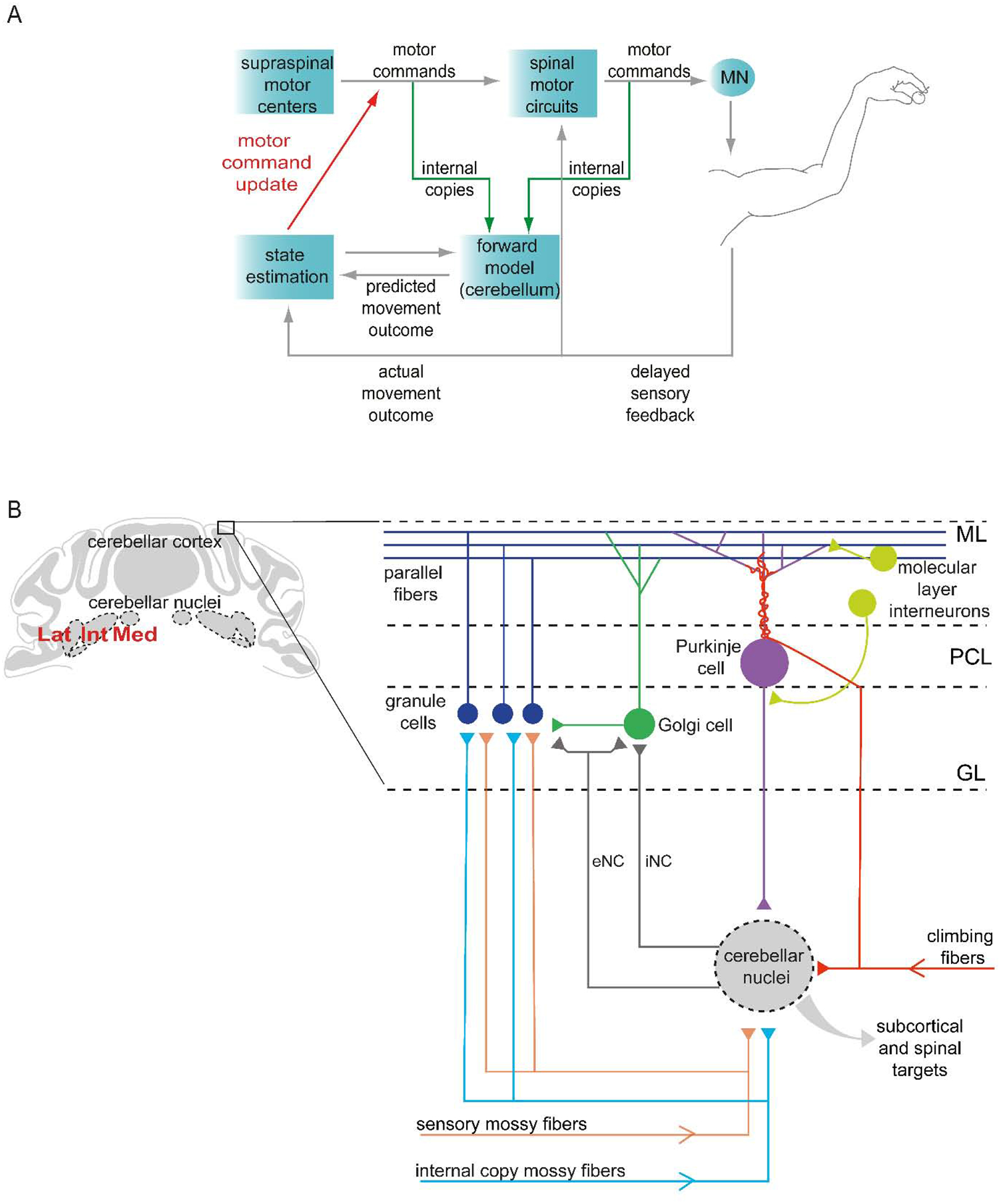
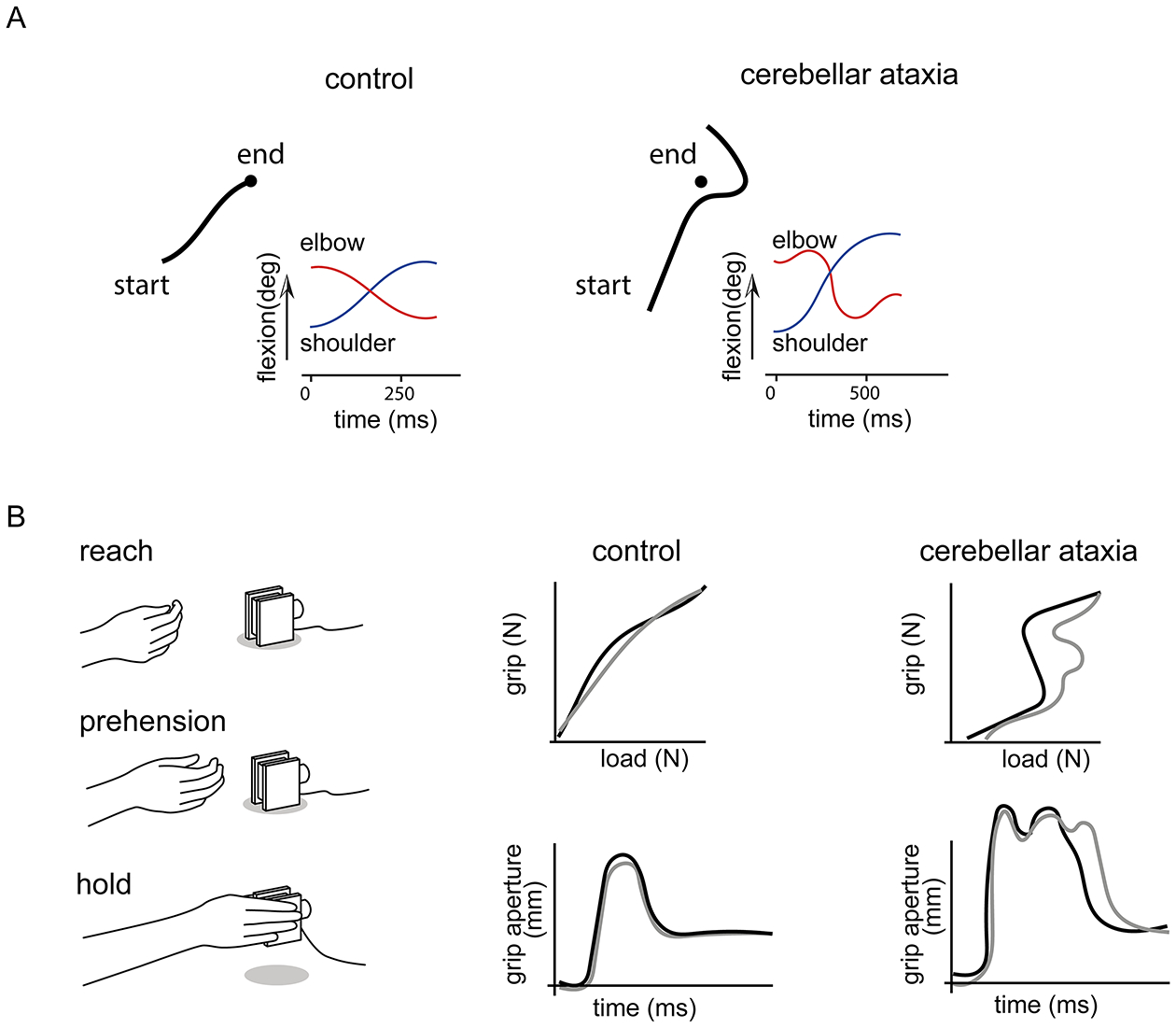
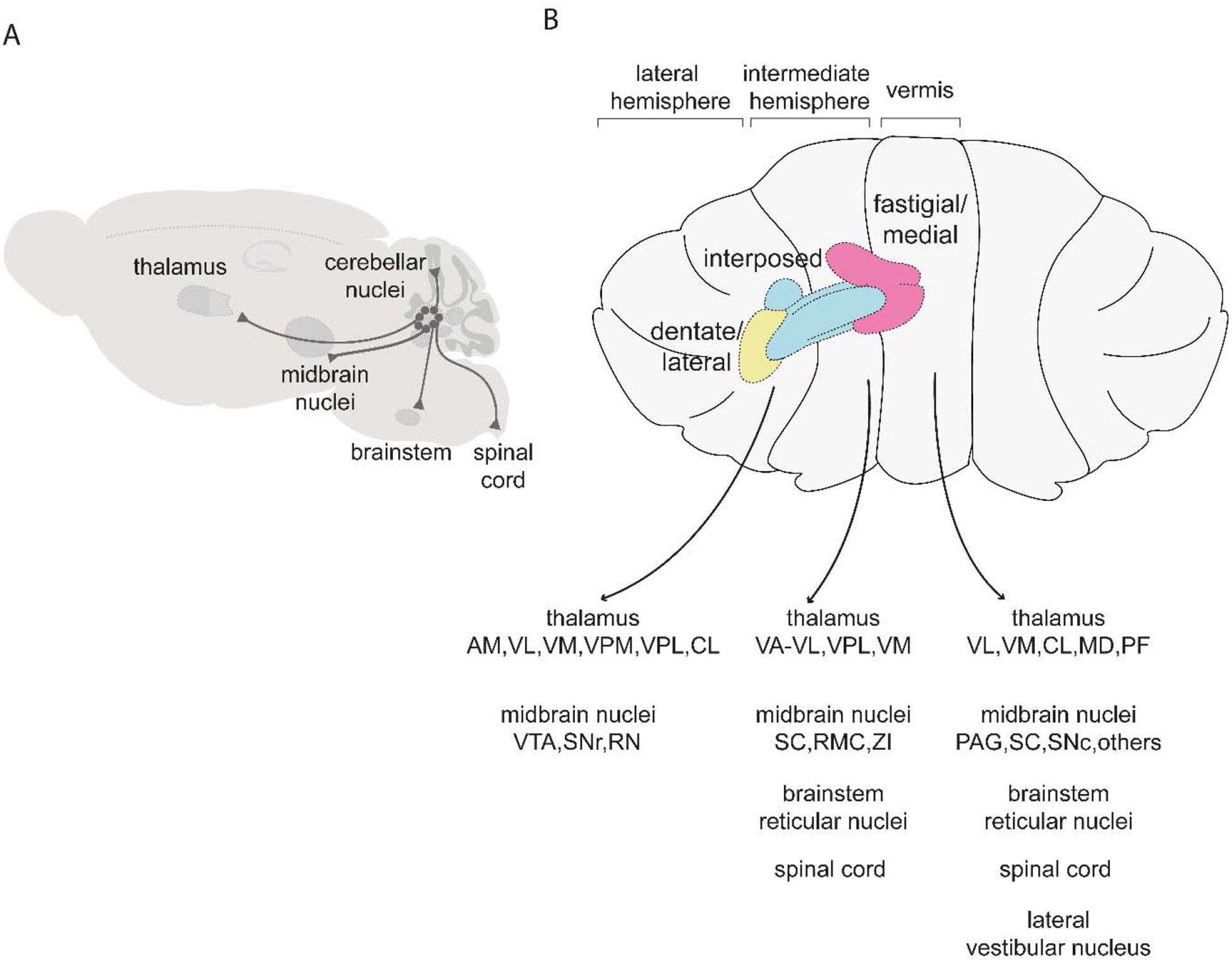
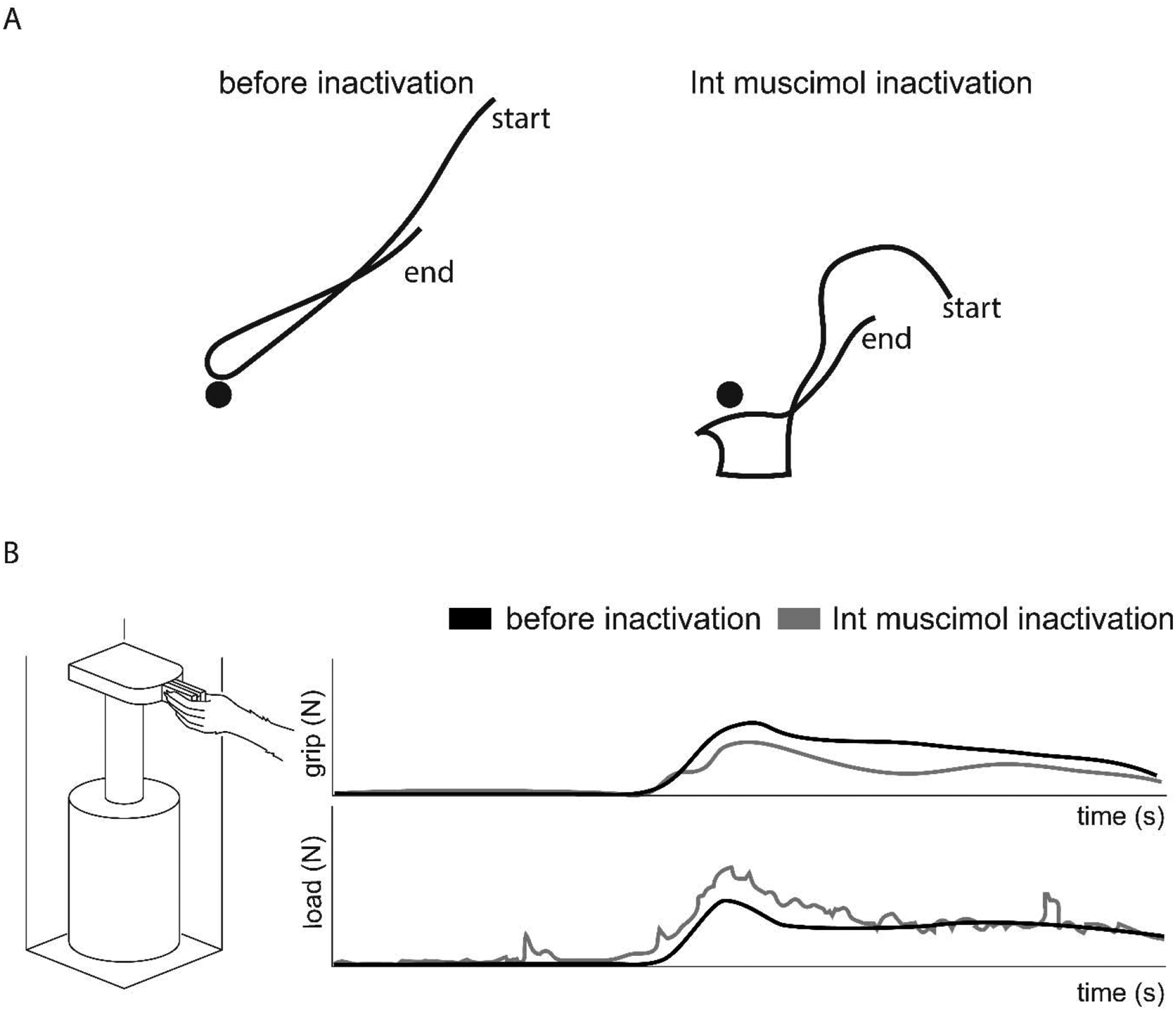
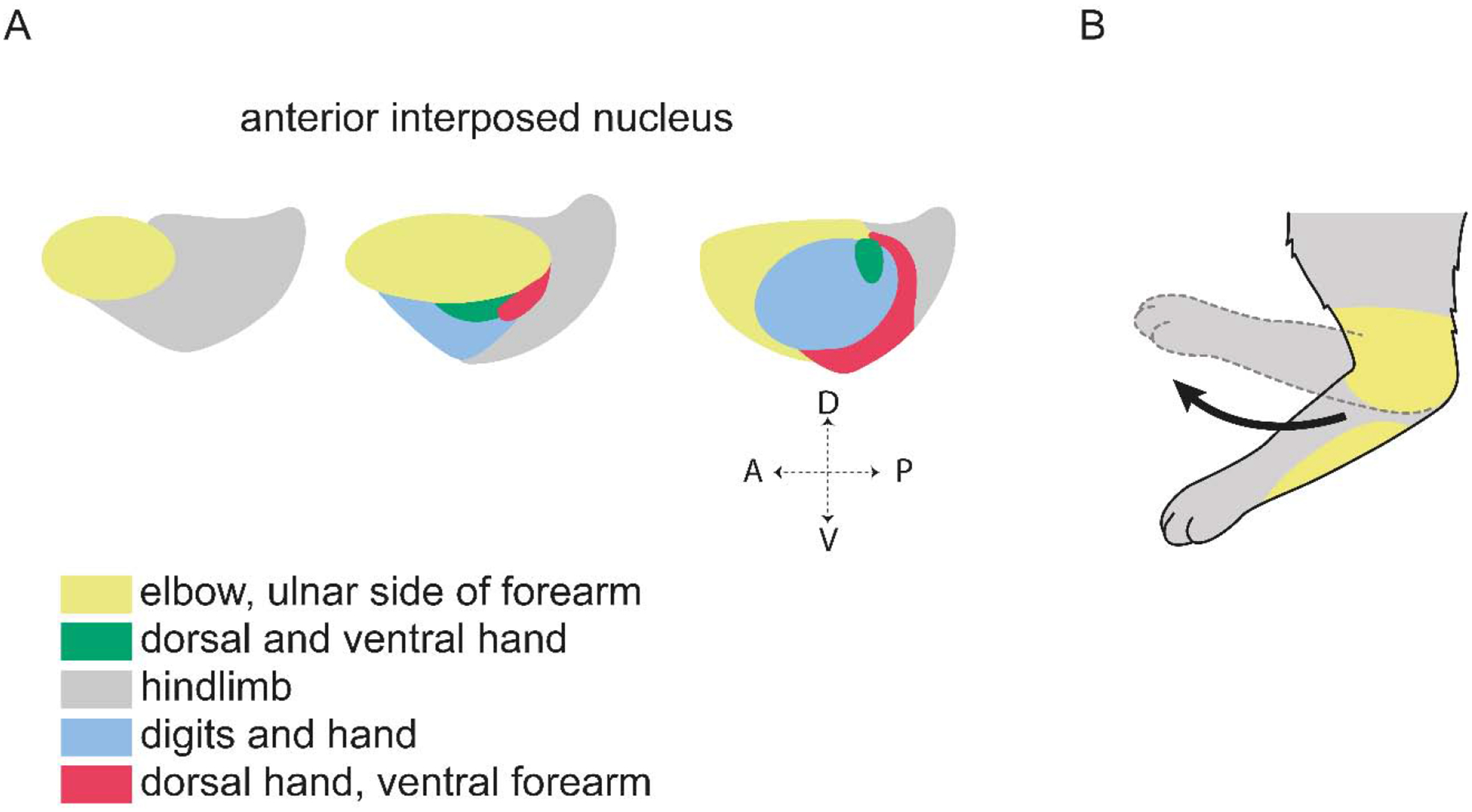
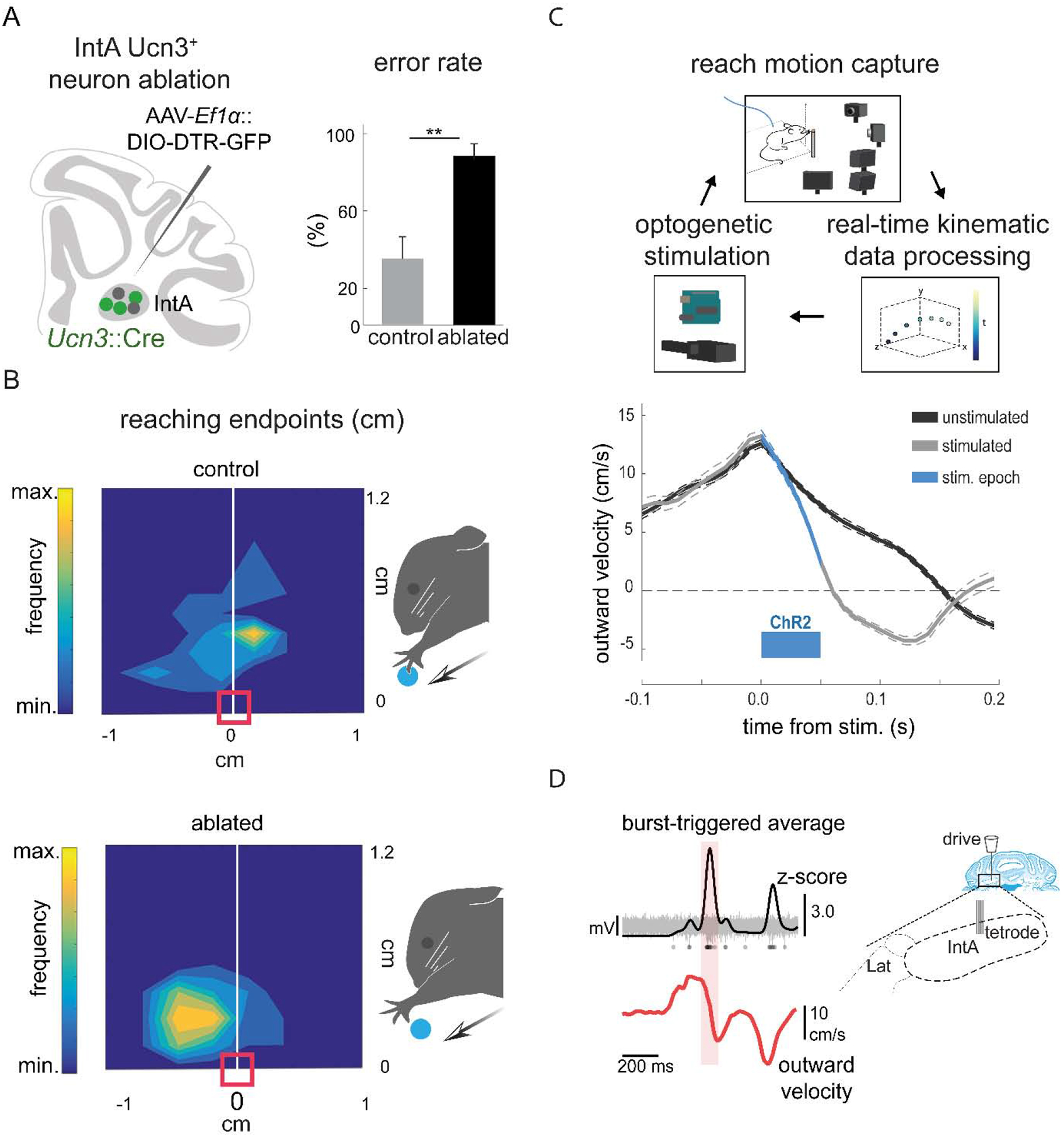
Dexterous forelimb movements like reaching, grasping, and manipulating objects are fundamental building blocks of the mammalian motor repertoire. These behaviors are essential to everyday activities, and their elaboration underlies incredible accomplishments by human beings in art and sport. Moreover, the susceptibility of these behaviors to damage and disease of the nervous system can lead to debilitating deficits, highlighting a need for a better understanding of function and dysfunction in sensorimotor control. The cerebellum is central to coordinating limb movements, as defined in large part by Joseph Babinski and Gordon Holmes describing motor impairment in patients with cerebellar lesions over 100 years ago (Babinski, 1902; Holmes, 1917), and supported by many important human and animal studies that have been conducted since. Here, with a focus on output pathways of the cerebellar nuclei across mammalian species, we describe forelimb movement deficits observed when cerebellar circuits are perturbed, the mechanisms through which these circuits influence motor output, and key challenges in defining how the cerebellum refines limb movement.
Keywords: Ataxia; Cerebellar nuclei; Dysmetria; Grasp; Internal copy; Reach.
Copyright © 2020 IBRO. Published by Elsevier Ltd. All rights reserved.
Figures
References
-
- Adrian MH, and John WK (2012). Theoretical Models of Motor Control and Motor Learning In Routledge Handbook of Motor Control and Motor Learning (Routledge; ).
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
