

Effects of ASC Application on Endplate Regeneration Upon Glycerol-Induced Muscle Damage
- PMID: 32655366
- PMCID: PMC7324987
- DOI: 10.3389/fnmol.2020.00107
Effects of ASC Application on Endplate Regeneration Upon Glycerol-Induced Muscle Damage
Abstract
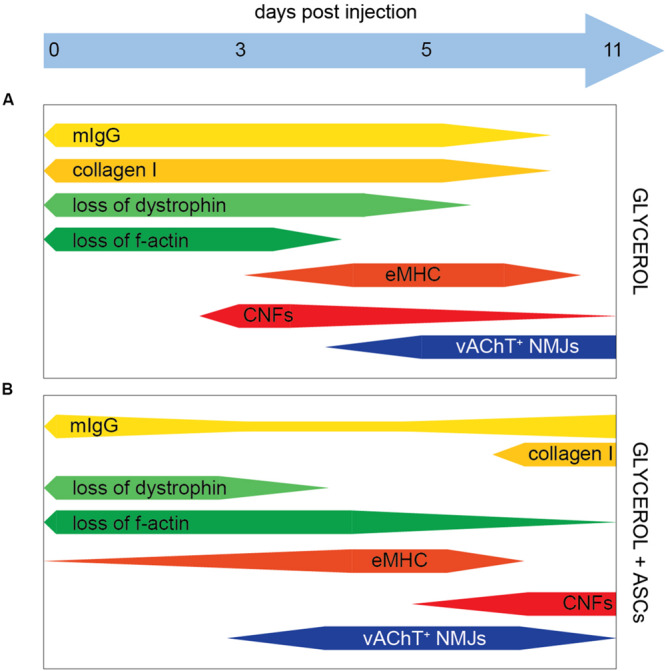
Amongst other approaches, adipose-derived stromal cells (ASCs) have recently been tested with respect to their regenerative capacity for treatment of neuromuscular disorders. While beneficial effects of ASCs on muscle recovery were observed previously, their impact on regeneration of neuromuscular junctions (NMJs) is unclear. Here, we used a murine glycerol damage model to study disruption and regeneration of NMJs and to evaluate the effects of systemic application of ASCs on muscle and NMJ recovery. In mice that were not treated with ASCs, a differential response of NMJ pre- and post-synapses to glycerol-induced damage was observed. While post-synapses were still present in regions that were necrotic and lacking actin and dystrophin, pre-synapses disappeared soon in those affected areas. Partial regeneration of NMJs occurred within 11 days after damage. ASC treatment slightly enhanced NMJ recovery and reduced the loss of presynaptic sites, but also led to a late phase of muscle necrosis and fibrosis. In summary, the results suggest a differential sensitivity of NMJ pre- and post-synapses to glycerol-induced muscle damage and that the use of ASC for the treatment of neuromuscular disorders needs further careful evaluation.
Keywords: ASC; NMJ; glycerol; regeneration; skeletal muscle.
Copyright © 2020 Rigon, Hörner, Straka, Bieback, Gretz, Hafner and Rudolf.
Figures




References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous