

CB1 Cannabinoid Receptors Stimulate Gβγ-GRK2-Mediated FAK Phosphorylation at Tyrosine 925 to Regulate ERK Activation Involving Neuronal Focal Adhesions
- PMID: 32655375
- PMCID: PMC7324865
- DOI: 10.3389/fncel.2020.00176
CB1 Cannabinoid Receptors Stimulate Gβγ-GRK2-Mediated FAK Phosphorylation at Tyrosine 925 to Regulate ERK Activation Involving Neuronal Focal Adhesions
Abstract
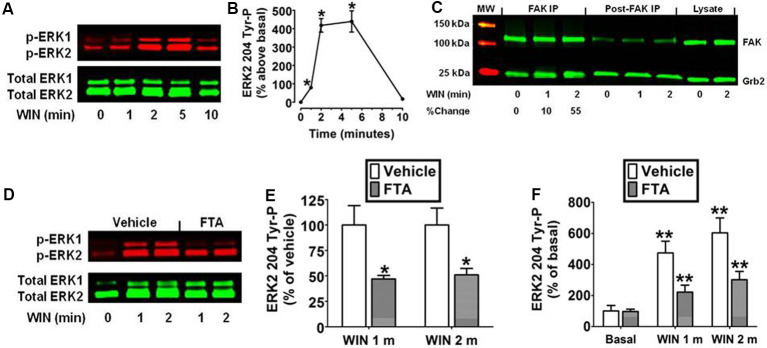
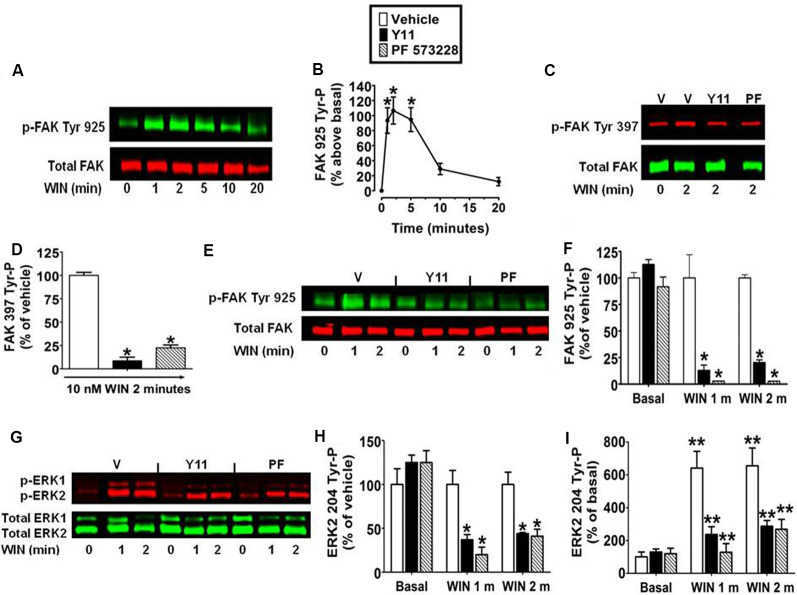
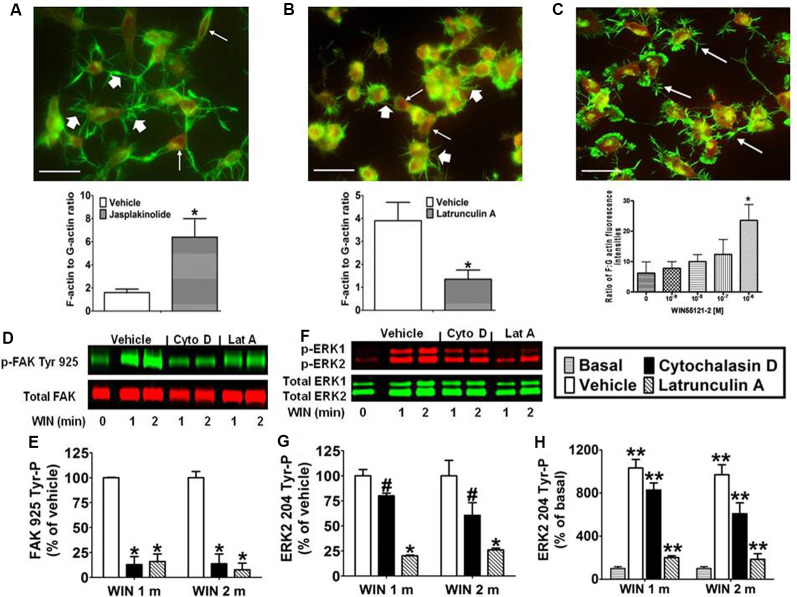
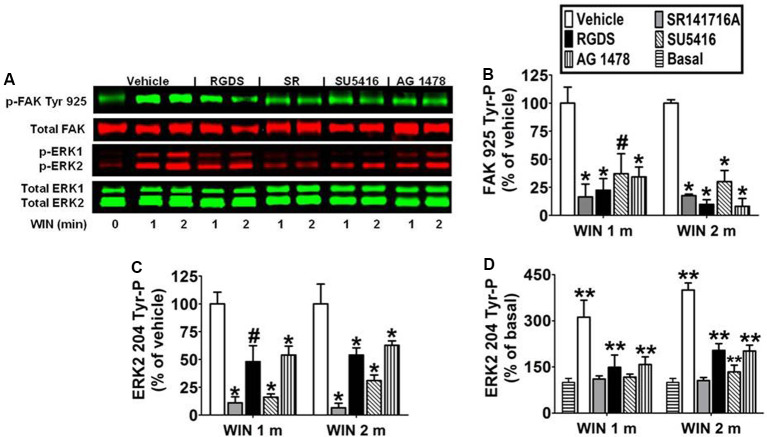
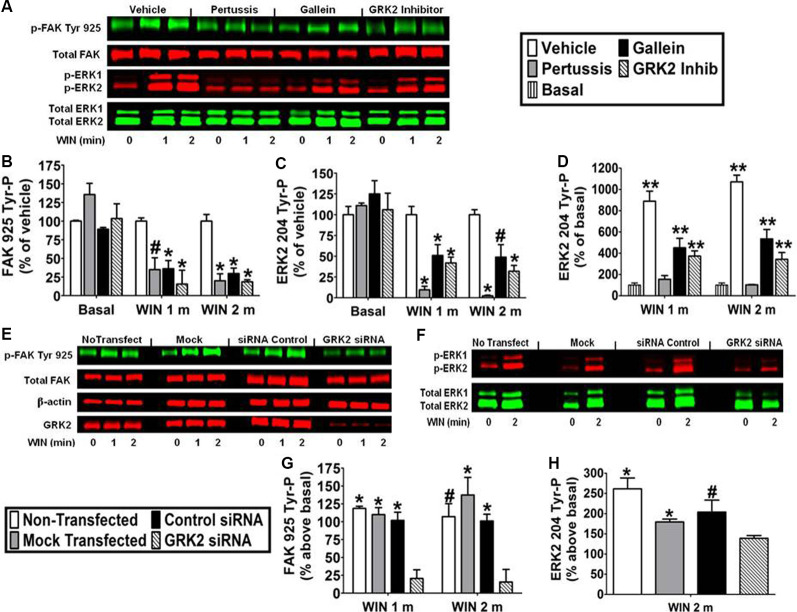
CB1 cannabinoid receptors (CB1) are abundantly expressed in the nervous system where they regulate focal adhesion kinase (FAK) and the mitogen-activated protein kinases (MAPK) extracellular signal-regulated kinase 1 and 2 (ERK1/2). However, the role of CB1-stimulated FAK 925 tyrosine phosphorylation (Tyr-P) in regulating ERK1/2 activation remains undefined. Here, immunoblotting analyses using antibodies against FAK phospho-Tyr 925 and ERK2 phospho-Tyr 204 demonstrated CB1-stimulated FAK 925 Tyr-P and ERK2 204 Tyr-P (0-5 min) which was followed by a decline in Tyr-P (5-20 min). CB1 stimulated FAK-Grb2 association and Ras-mediated ERK2 activation. The FAK inhibitors Y11 and PF 573228 abolished FAK 925 Tyr-P and partially inhibited ERK2 204 Tyr-P. FAK 925 Tyr-P and ERK2 204 Tyr-P were adhesion-dependent, required an intact actin cytoskeleton, and were mediated by integrins, Flk-1 vascular endothelial growth factor receptors, and epidermal growth factor receptors. FAK 925 Tyr-P and ERK2 204 Tyr-P were blocked by the Gβγ inhibitor gallein, a GRK2 inhibitor, and GRK2 siRNA silencing, suggesting Gβγ and GRK2 participate in FAK-mediated ERK2 activation. Together, these studies indicate FAK 925 Tyr-P occurs concurrently with CB1-stimulated ERK2 activation and requires the actin cytoskeleton and Gi/oβγ-GRK2-mediated cross-talk between CB1, integrins, and receptor tyrosine kinases (RTKs).
Keywords: CB1; ERK; FAK; GRK2; Grb2; Gβγ.
Copyright © 2020 Dalton, Carney, Marshburn, Norford and Howlett.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous