

Internalization and Intoxication of Human Macrophages by the Active Subunit of the Aggregatibacter actinomycetemcomitans Cytolethal Distending Toxin Is Dependent Upon Cellugyrin (Synaptogyrin-2)
- PMID: 32655562
- PMCID: PMC7325893
- DOI: 10.3389/fimmu.2020.01262
Internalization and Intoxication of Human Macrophages by the Active Subunit of the Aggregatibacter actinomycetemcomitans Cytolethal Distending Toxin Is Dependent Upon Cellugyrin (Synaptogyrin-2)
Abstract
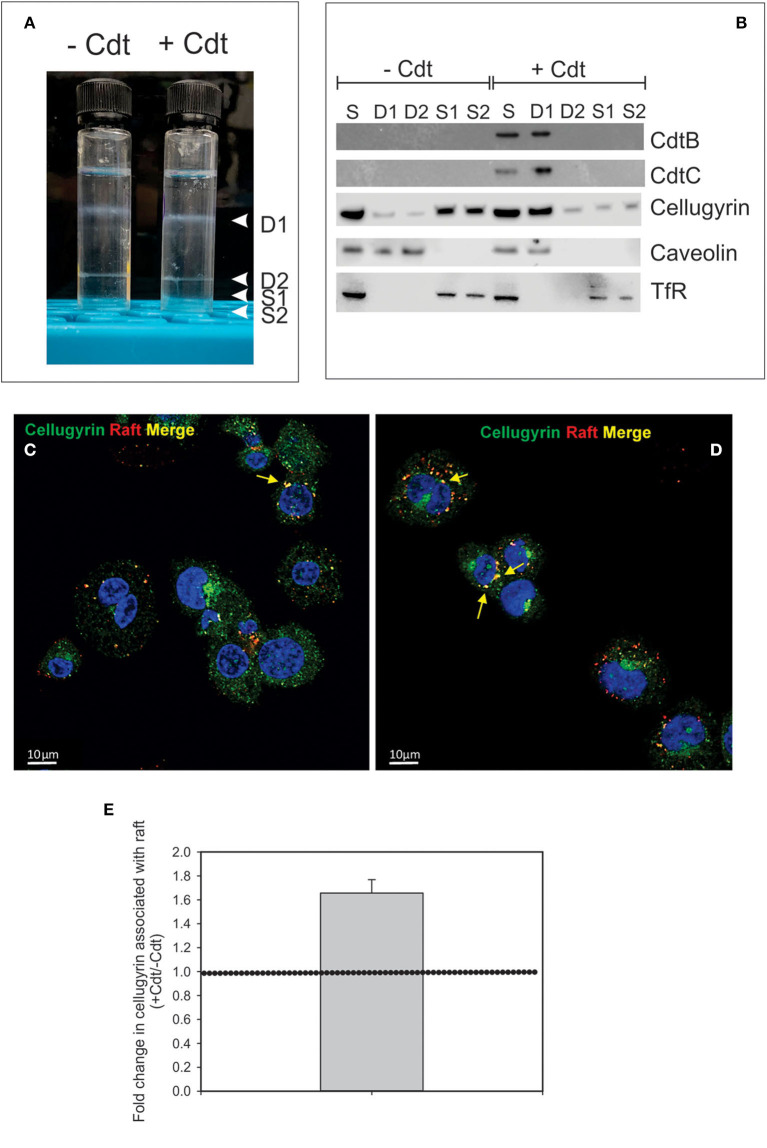
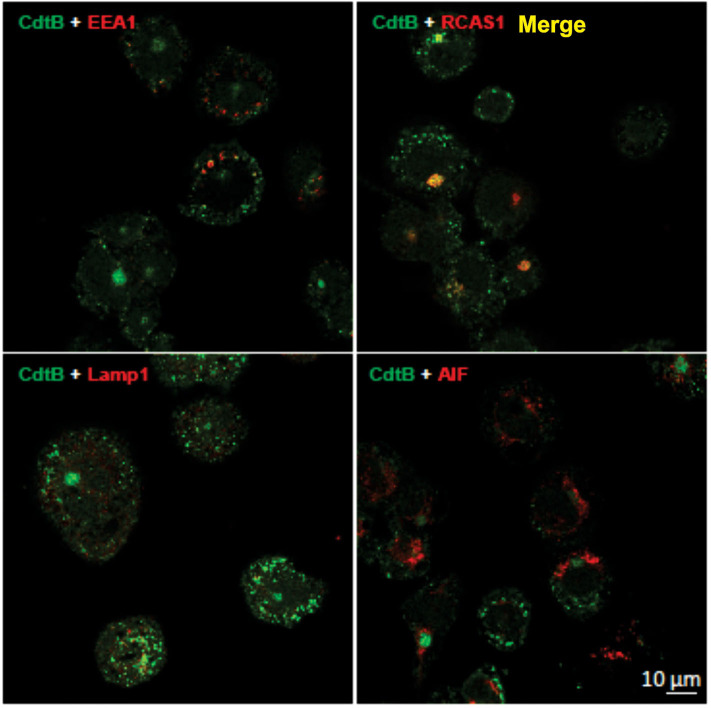
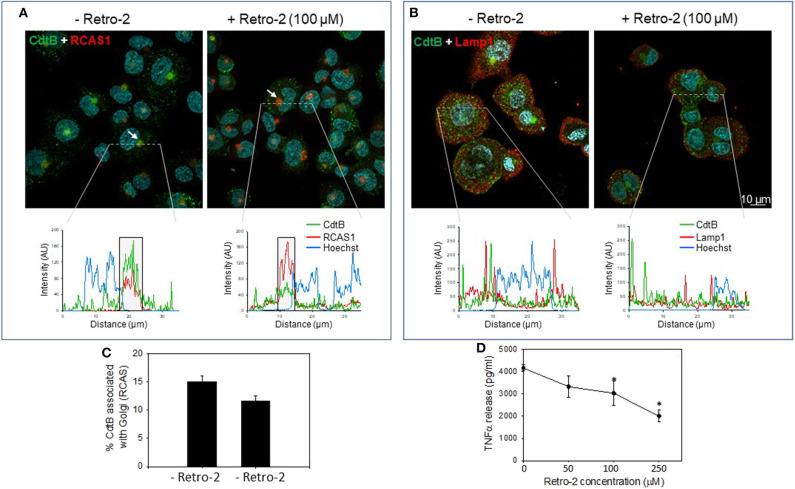
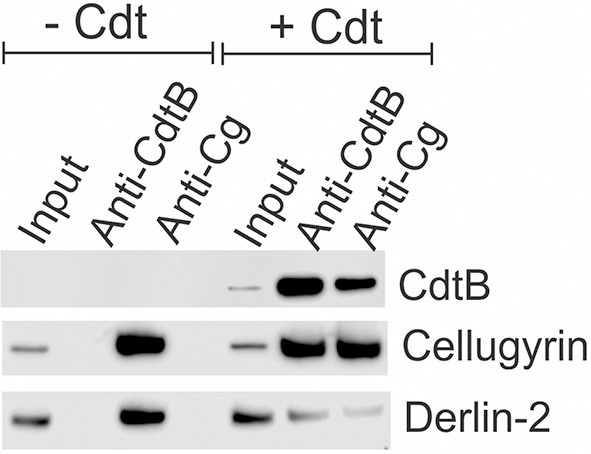
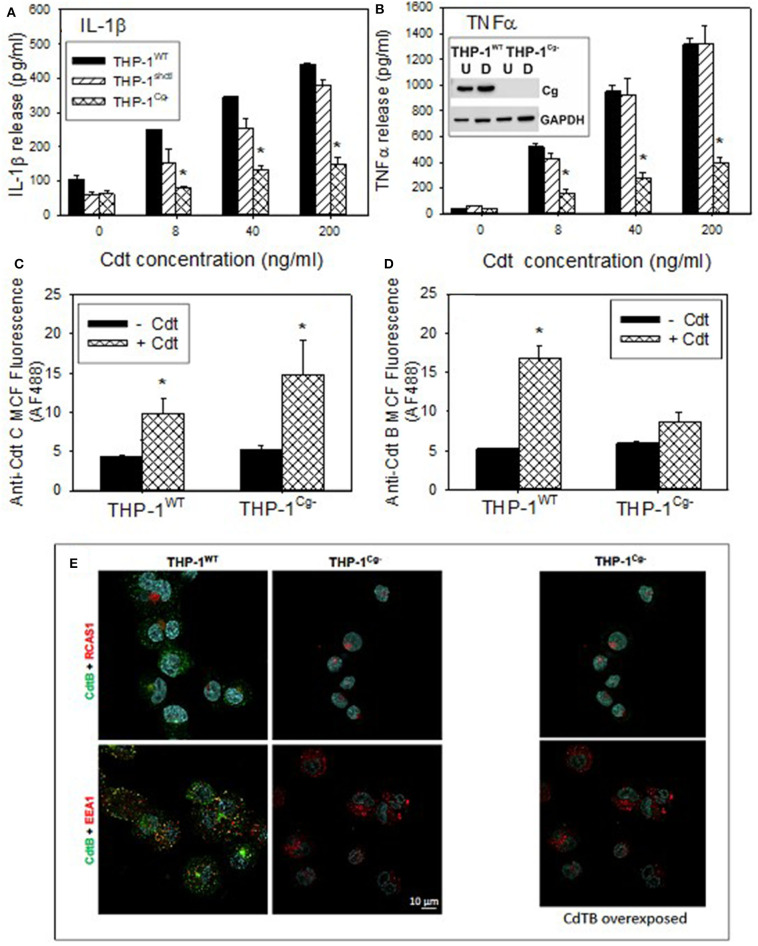
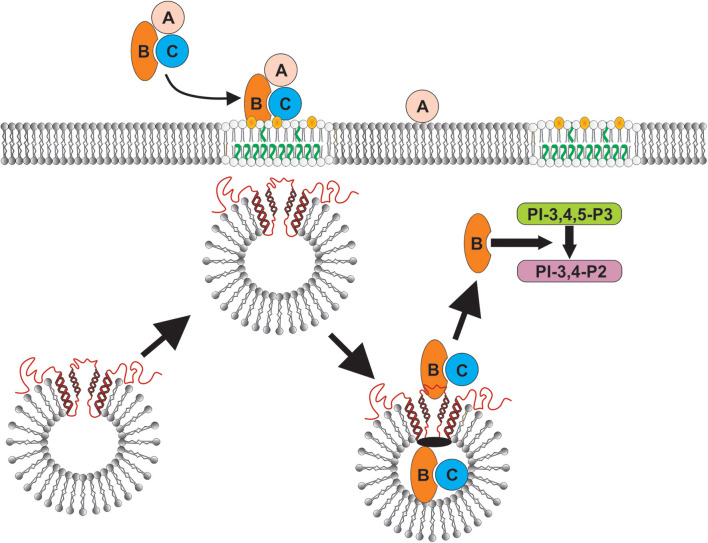
The Aggregatibacter actinomycetemcomitans cytolethal distending toxin (Cdt) is a heterotrimeric AB2 toxin capable of inducing cell cycle arrest and apoptosis in lymphocytes and other cell types. Recently, we have demonstrated that human macrophages are resistant to Cdt-induced apoptosis but are susceptible to toxin-induced pro-inflammatory cytokine response involving activation of the NLRP3 inflammasome. Exposure to Cdt results in binding to the cell surface followed by internalization and translocation of the active subunit, CdtB, to intracellular compartments. Internalization involves hijacking of retrograde pathways; treatment of cells with Retro-2 leads to a decrease in CdtB-Golgi association. These events are dependent upon toxin binding to cholesterol in the context of lipid rich membrane microdomains often referred to as lipid rafts. We now demonstrate that within 1 h of exposure of macrophages to Cdt, CdtB is internalized and found primarily within lipid rafts; concurrently, cellugyrin (synaptogyrin-2) also translocates into lipid rafts. Further analysis by immunoprecipitation indicates that CdtB associates with complexes containing both cellugyrin and Derlin-2. Moreover, a human macrophage cell line deficient in cellugyrin expression (THP-1Cg-) challenged with Cdt failed to internalize CdtB and was resistant to the Cdt-induced pro-inflammatory response. We propose that lipid rafts along with cellugyrin play a critical role in the internalization and translocation of CdtB to critical intracellular target sites in human macrophages. These studies provide the first evidence that cellugyrin is expressed in human macrophages and plays a critical role in Cdt toxicity of these cells.
Keywords: Aggregatibacter actinomycetemcomitans; bacterial toxins; cellugyrin; cytolethal distending toxin; lipid rafts; macrophages.
Copyright © 2020 Boesze-Battaglia, Dhingra, Walker, Zekavat and Shenker.
Figures
References
-
- Fox JG, Rogers AB, Whary MT, Ge Z, Taylor NS, Xu S, et al. Gastroenteritis in NF-kappaB-deficient mice is produced with wild-type Camplyobacter jejuni but not with C. jejuni lacking cytolethal distending toxin despite persistent colonization with both strains. Infect Immun. (2004) 72:1116–25. 10.1128/iai.72.2.1116-1125.2004 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources