

Effects of Potassium Levels on Plant Growth, Accumulation and Distribution of Carbon, and Nitrate Metabolism in Apple Dwarf Rootstock Seedlings
- PMID: 32655607
- PMCID: PMC7325393
- DOI: 10.3389/fpls.2020.00904
Effects of Potassium Levels on Plant Growth, Accumulation and Distribution of Carbon, and Nitrate Metabolism in Apple Dwarf Rootstock Seedlings
Abstract
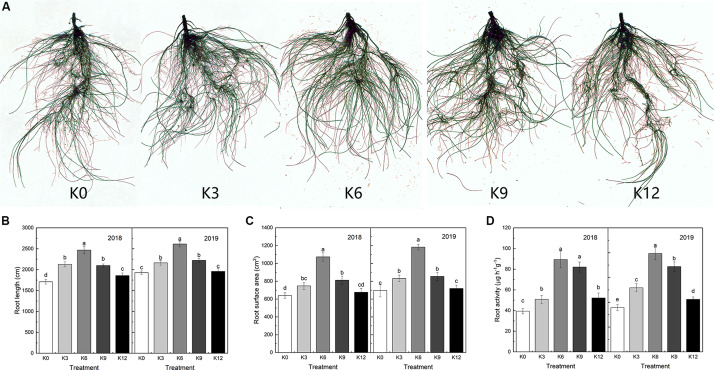
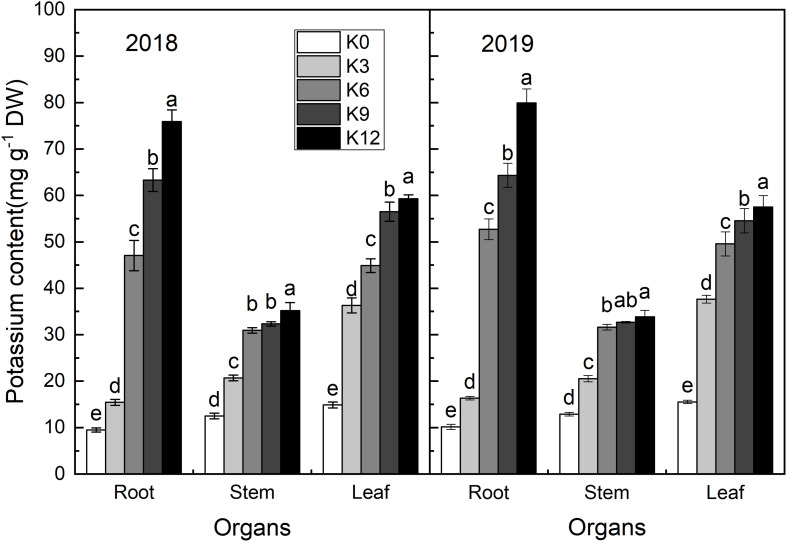
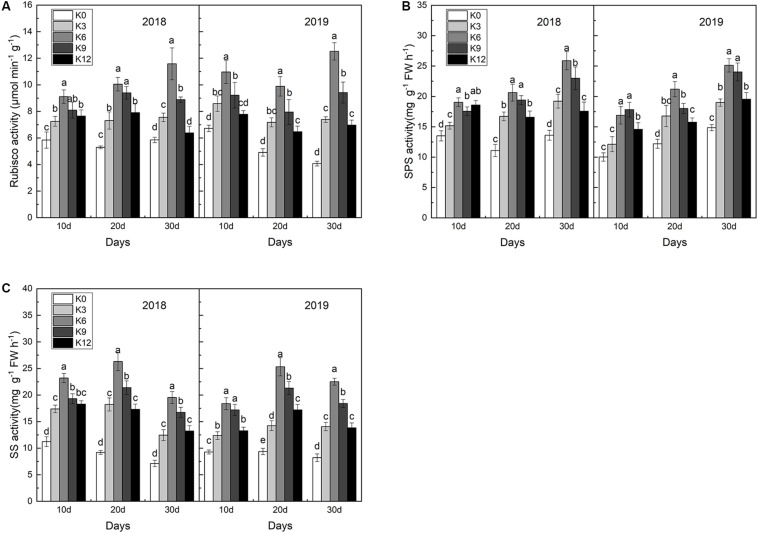
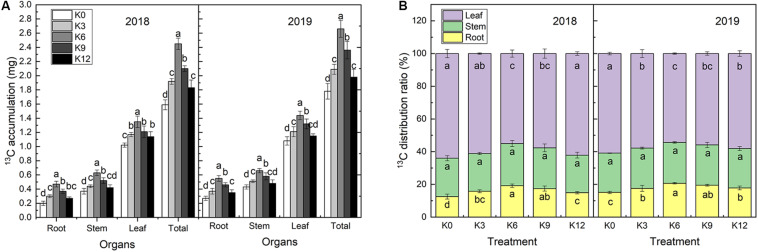
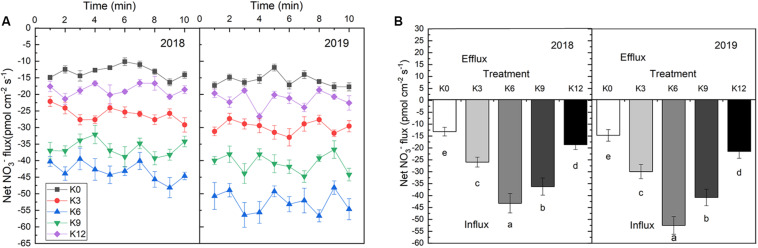
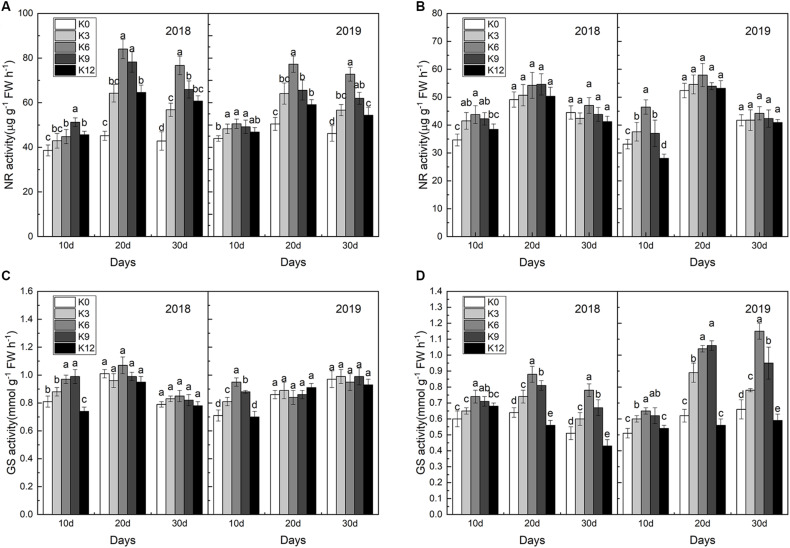
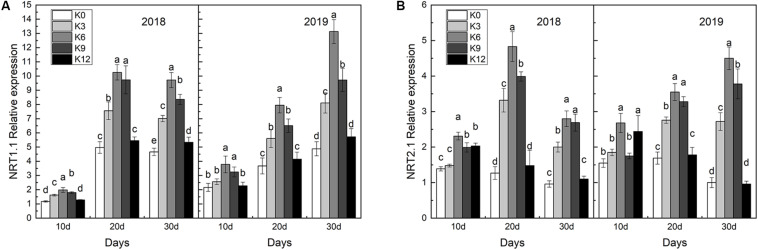
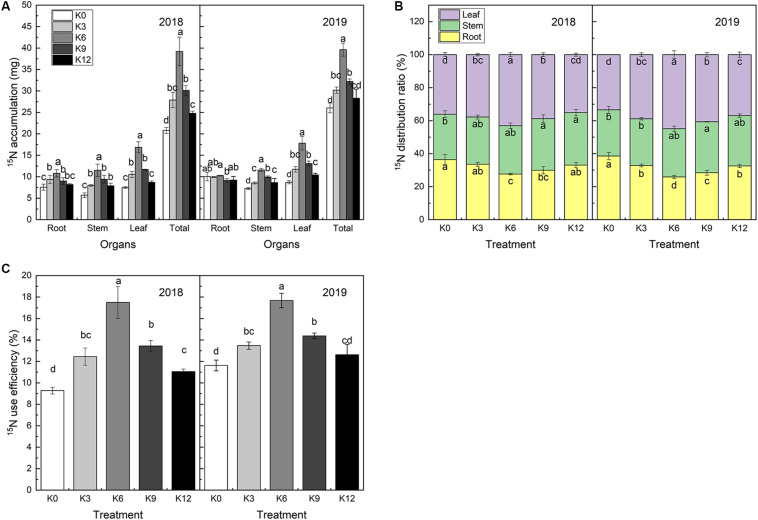
Nitrogen (N) is one of the most required mineral elements for plant growth, and potassium (K) plays a vital role in nitrogen metabolism, both elements being widely applied as fertilizers in agricultural production. However, the exact relationship between K and nitrogen use efficiency (NUE) remains unclear. Apple dwarf rootstock seedlings (M9T337) were used to study the impacts of different K levels on plant growth, nitrogen metabolism, and carbon (C) assimilation in water culture experiments for 2 years. The results showed that both deficiency and excess K inhibited the growth and root development of M9T337 seedlings. When the K supply concentration was 0 mM and 12 mM, the biomass of each organ, root-shoot ratio, root activity and NO3 - ion flow rate decreased significantly, net photosynthetic rate (P n) and photochemical efficiency (F v/F m) being lower. Meanwhile, seedlings treated with 6 mM K+ had higher N and C metabolizing enzyme activities and higher nitrate transporter gene expression levels (NRT1.1; NRT2.1). 13C and 15N labeling results showed that deficiency and excess K could not only reduce 15N absorption and 13C assimilation accumulation of M9T337 seedlings, but also reduced the 15N distribution ratio in leaves and 13C distribution ratio in roots. These results suggest that appropriate K supply (6 mM) was optimal as it enhanced photoassimilate transport from leaves to roots and increased NUE by influencing photosynthesis, C and N metabolizing enzyme activities, nitrate assimilation gene activities, and nitrate transport.
Keywords: 13C; 15N; M9T337 seedlings; NUE; biomass; enzyme activity; nitrate transporter gene expression.
Copyright © 2020 Xu, Du, Wang, Sha, Chen, Tian, Zhu, Ge and Jiang.
Figures
References
-
- Armengaud P., Sulpice R., Miller A. J., Stitt M., Amtmann A., Gibon Y. (2009). Multilevel analysis of primary metabolism provides new insights into the role of potassium nutrition for glycolysis and nitrogen assimilation in Arabidopsis roots. Plant Physiol. 150 772–785. 10.1104/pp.108.133629 - DOI - PMC - PubMed
-
- Ben Zioni A., Vaadia Y., Herman L. S. (1971). Nitrate uptake by roots as regulated by nitrate reduction products of the shoot. Physiol. Plantarum. 24 288–290. 10.1111/j.1399-3054.1971.tb03493.x - DOI
LinkOut - more resources
Full Text Sources