

The role of lipids in autophagy and its implication in neurodegeneration
- PMID: 32656499
- PMCID: PMC7328674
- DOI: 10.15698/cst2020.07.225
The role of lipids in autophagy and its implication in neurodegeneration
Abstract
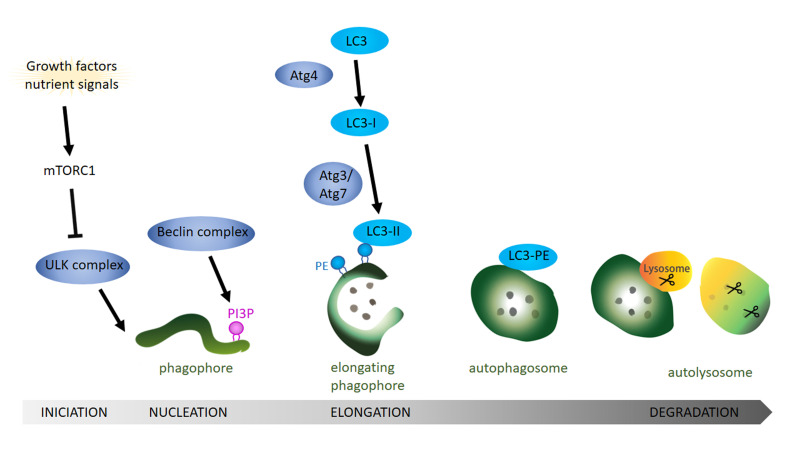
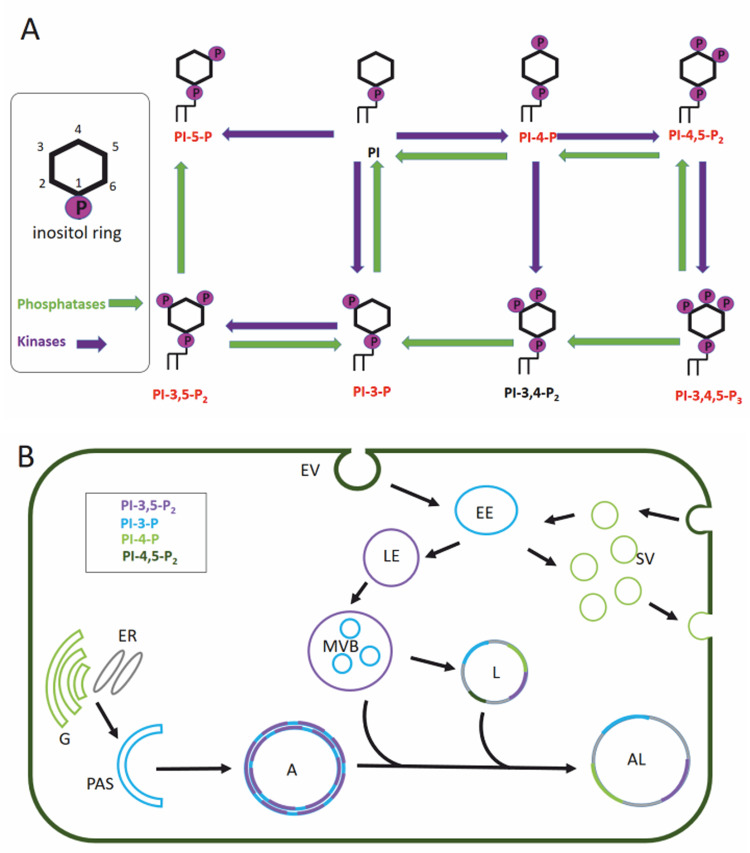
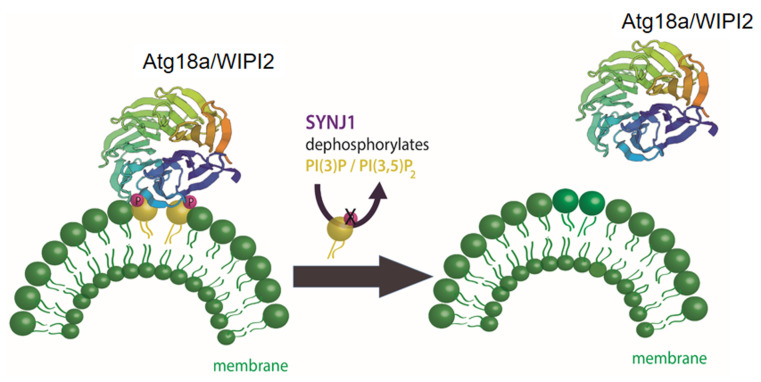
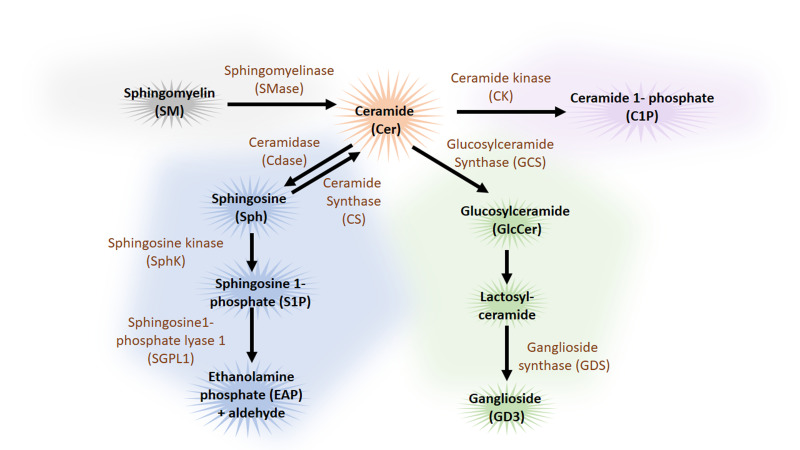
Neurodegenerative diseases are, at present, major socio-economic burdens without effective treatments and their increasing prevalence means that these diseases will be a challenge for future generations. Neurodegenerative diseases may differ in etiology and pathology but are often caused by the accumulation of dysfunctional and aggregation-prone proteins. Autophagy, a conserved cellular mechanism, deals with cellular stress and waste product build-up and has been shown to reduce the accumulation of dysfunctional proteins in animal models of neurodegenerative diseases. Historically, progress in understanding the precise function of lipids has traditionally been far behind other biological molecules (like proteins) but emerging works demonstrate the importance of lipids in the autophagy pathway and how the disturbance of lipid metabolism is connected to neurodegeneration. Here we review how altered autophagy and the disturbance of lipid metabolism, particularly of phosphoinositols and sphingolipids, feature in neurodegenerative diseases and address work from the field that suggests that these potentially offer an opportunity of therapeutic intervention.
Keywords: Alzheimer's disease; Parkinson's disease; autophagy; lipids; neurodegeneration; phosphoinositols; sphingolipids.
Copyright: © 2020 Hernandez-Diaz and Soukup.
Conflict of interest statement
Conflict of interest: The authors declare no conflict of interest.
Figures


References
-
- Dorsey ER, Constantinescu R, Thompson JP, Biglan KM, Holloway RG, Kieburtz K, Marshall FJ, Ravina BM, Schifitto G, Siderowf A, Tanner CM. Projected number of people with Parkinson disease in the most populous nations, 2005 through 2030. Neurology. 2007;68:384–386. doi: 10.1212/01.wnl.0000271777.50910.73. - DOI - PubMed
Publication types
LinkOut - more resources
Full Text Sources