

FcRn is a CD32a coreceptor that determines susceptibility to IgG immune complex-driven autoimmunity
- PMID: 32658257
- PMCID: PMC7537387
- DOI: 10.1084/jem.20200359
FcRn is a CD32a coreceptor that determines susceptibility to IgG immune complex-driven autoimmunity
Abstract
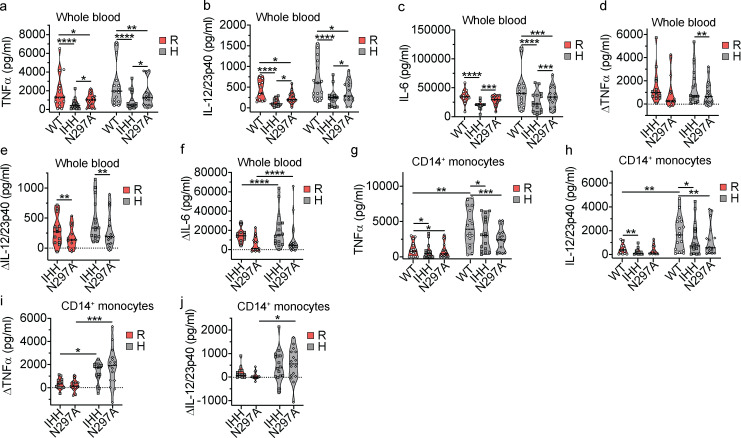
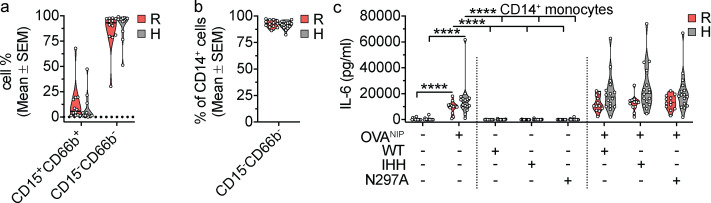
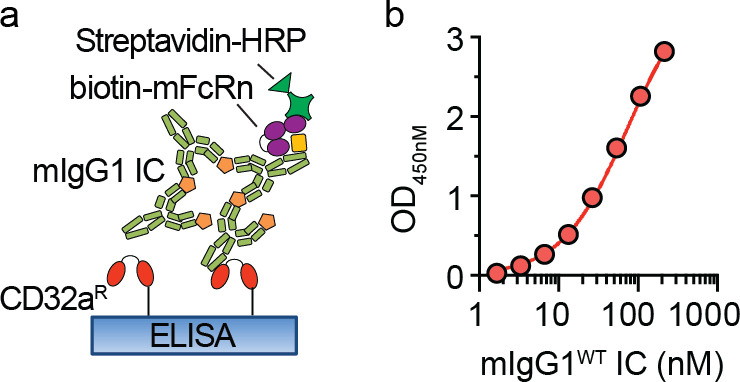
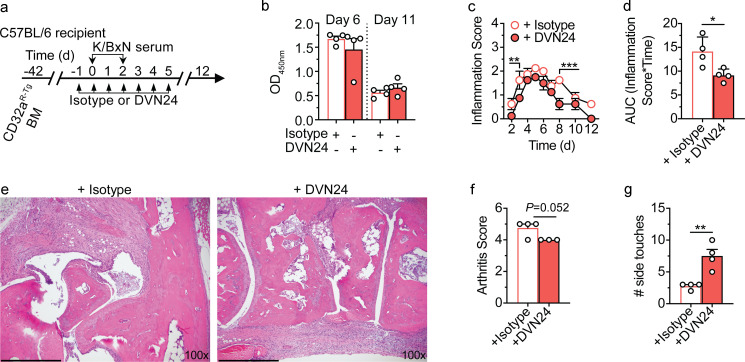
IgG immune complexes (ICs) promote autoimmunity through binding fragment crystallizable (Fc) γ-receptors (FcγRs). Of these, the highly prevalent FcγRIIa (CD32a) histidine (H)-131 variant (CD32aH) is strongly linked to human autoimmune diseases through unclear mechanisms. We show that, relative to the CD32a arginine (R)-131 (CD32aR) variant, CD32aH more avidly bound human (h) IgG1 IC and formed a ternary complex with the neonatal Fc receptor (FcRn) under acidic conditions. In primary human and mouse cells, both CD32a variants required FcRn to induce innate and adaptive immune responses to hIgG1 ICs, which were augmented in the setting of CD32aH. Conversely, FcRn induced responses to IgG IC independently of classical FcγR, but optimal responses required FcRn and FcγR. Finally, FcRn blockade decreased inflammation in a rheumatoid arthritis model without reducing circulating autoantibody levels, providing support for FcRn's direct role in IgG IC-associated inflammation. Thus, CD32a and FcRn coregulate IgG IC-mediated immunity in a manner favoring the CD32aH variant, providing a novel mechanism for its disease association.
© 2020 Hubbard et al.
Conflict of interest statement
Disclosures: J.J. Hubbard reported a patent to US2019/017880 pending. M. Pyzik reported a patent to PCT/US2019/017880 pending, "Brigham and Women's Hospital." T. Rath reported a patent to 61/984,652 licensed and a patent to 61/909,229 licensed. A.K. Gandhi reported a patent for therapeutic FcRn-based bispecific monoclonal antibodies pending (PCT/US2019/107880). D.C. Roopenian reported a patent to US20190135915A1 licensed, "Alexion Pharmaceuticals"; and served as consultant with equity interests in Syntimmune Inc., a company developing therapeutic agents to target FcRn. Syntimmune is now a wholly owned subsidiary of Alexion Pharmaceuticals, Inc., following its acquisition by Alexion in November 2018. I. Sandlie has served as a consultant with equity interests in Syntimmune Inc., a company developing therapeutic agents to target FcRn. Syntimmune is now a wholly owned subsidiary of Alexion Pharmaceuticals, Inc., following its acquisition by Alexion in November 2018. K. Baker reported a patent to 61/984,652 issued and a patent to 61/909,229 issued. R.S. Blumberg served as consultant with equity interests in Syntimmune Inc., a company developing therapeutic agents to target FcRn. Syntimmune is now a wholly owned subsidiary of Alexion Pharmaceuticals, Inc., following its acquisition by Alexion in November 2018. In addition, R.S. Blumberg has a pending patent PCT/US2019/017880 to “BWH" and two patents licensed to "Alexion" (US2017/002073 and US2017/0045528). No other disclosures were reported.
Figures
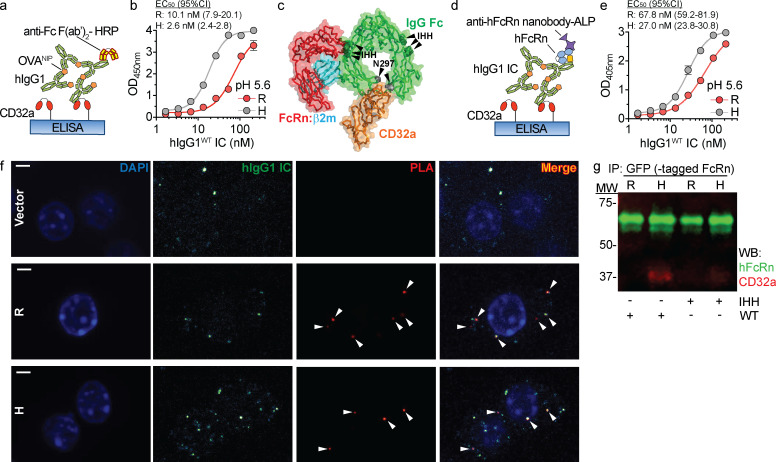
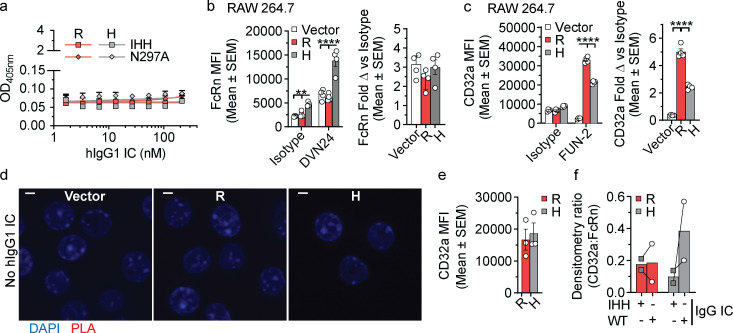
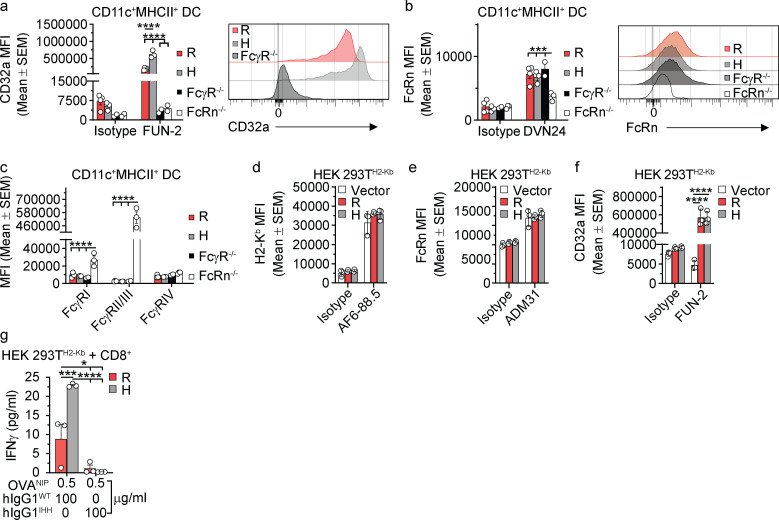
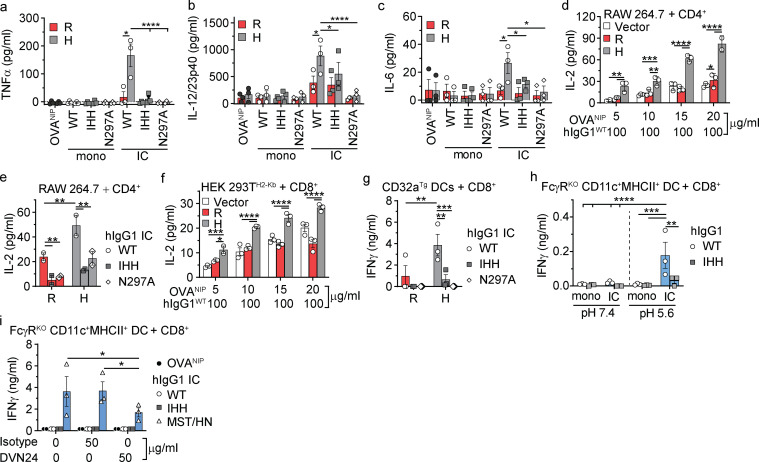
References
-
- Baker K., Qiao S.W., Kuo T.T., Aveson V.G., Platzer B., Andersen J.T., Sandlie I., Chen Z., de Haar C., Lencer W.I., et al. . 2011. Neonatal Fc receptor for IgG (FcRn) regulates cross-presentation of IgG immune complexes by CD8-CD11b+ dendritic cells. Proc. Natl. Acad. Sci. USA. 108:9927–9932. 10.1073/pnas.1019037108 - DOI - PMC - PubMed
-
- Baker K., Rath T., Flak M.B., Arthur J.C., Chen Z., Glickman J.N., Zlobec I., Karamitopoulou E., Stachler M.D., Odze R.D., et al. . 2013. Neonatal Fc receptor expression in dendritic cells mediates protective immunity against colorectal cancer. Immunity. 39:1095–1107. 10.1016/j.immuni.2013.11.003 - DOI - PMC - PubMed
-
- Blumberg L.J., Humphries J.E., Jones S.D., Pearce L.B., Holgate R., Hearn A., Cheung J., Mahmood A., Del Tito B., Graydon J.S., et al. . 2019. Blocking FcRn in humans reduces circulating IgG levels and inhibits IgG immune complex-mediated immune responses. Sci. Adv. 5:x9586 10.1126/sciadv.aax9586 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
