

A Role of Epithelial Cells and Virulence Factors in Biofilm Formation by Streptococcus pyogenes In Vitro
- PMID: 32661124
- PMCID: PMC7504962
- DOI: 10.1128/IAI.00133-20
A Role of Epithelial Cells and Virulence Factors in Biofilm Formation by Streptococcus pyogenes In Vitro
Abstract
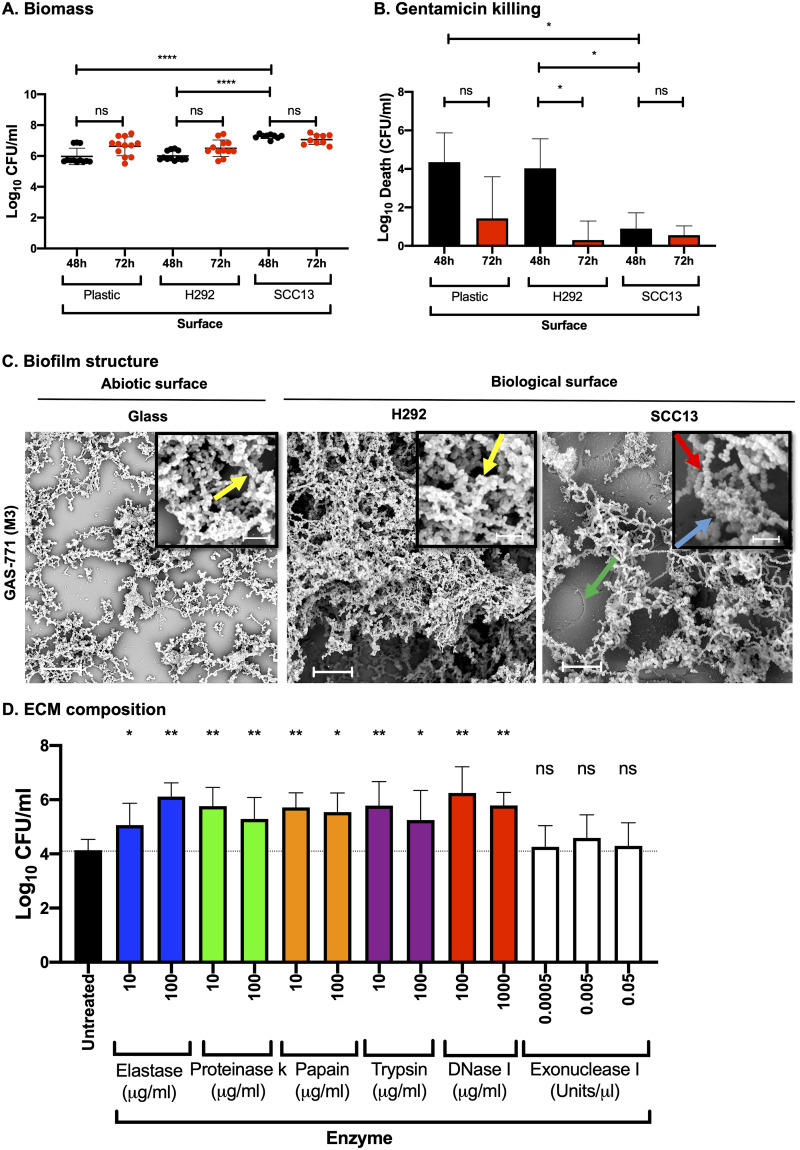
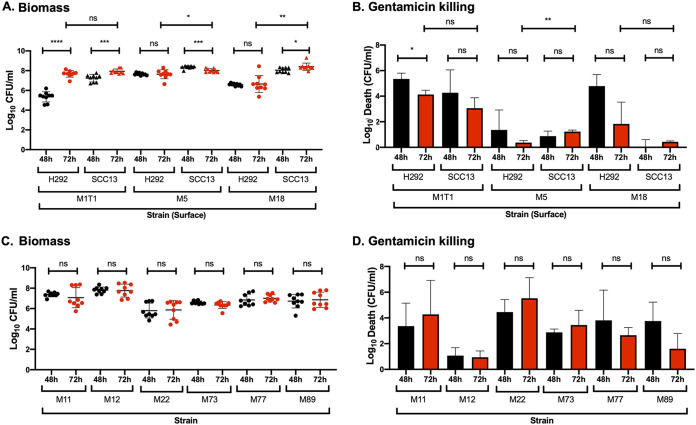
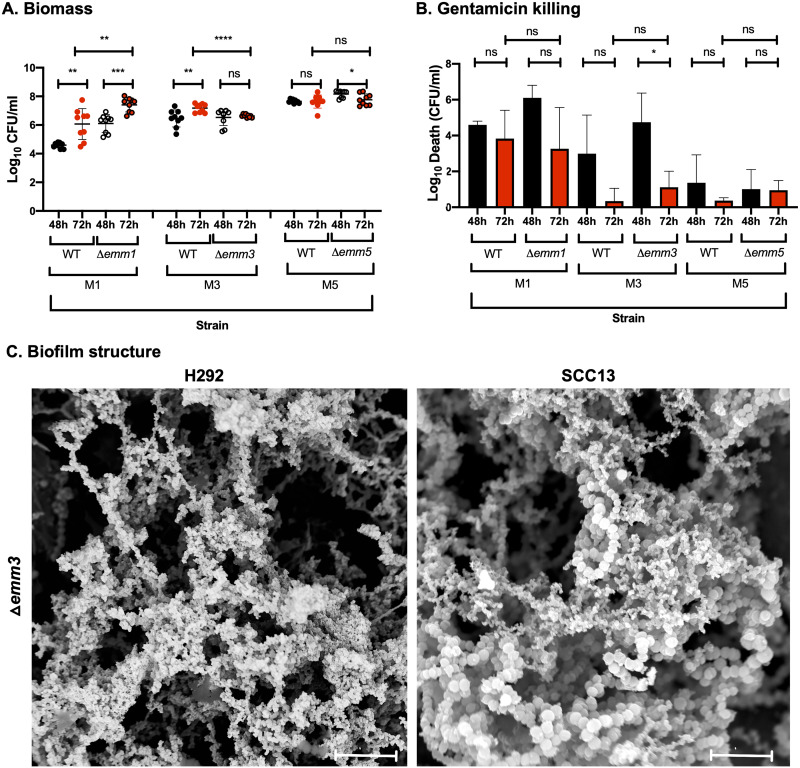
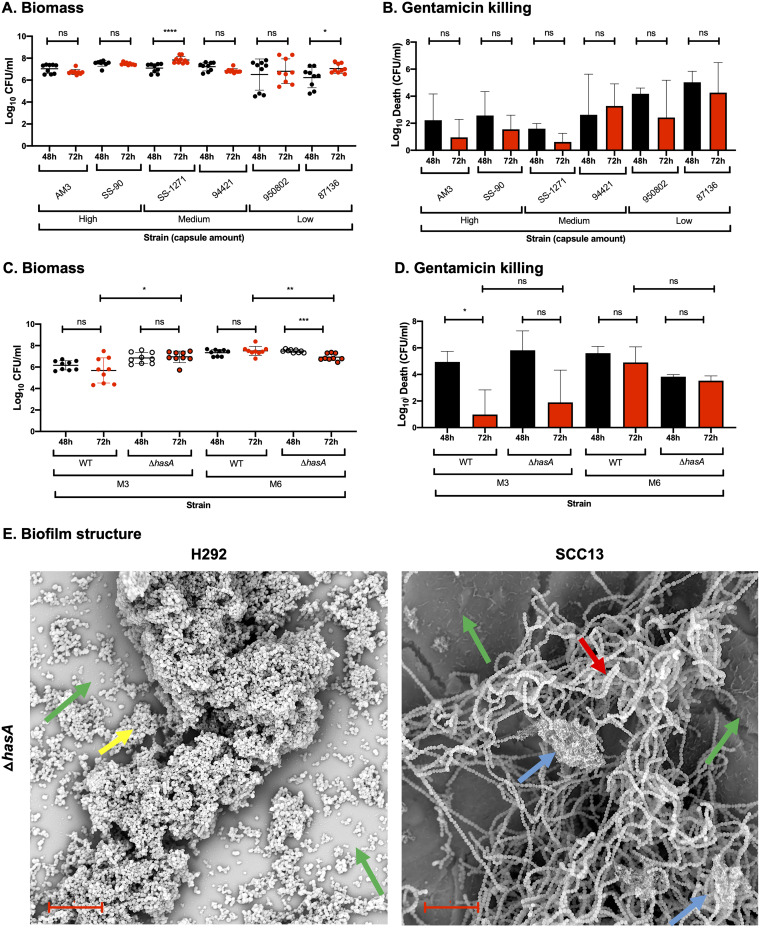
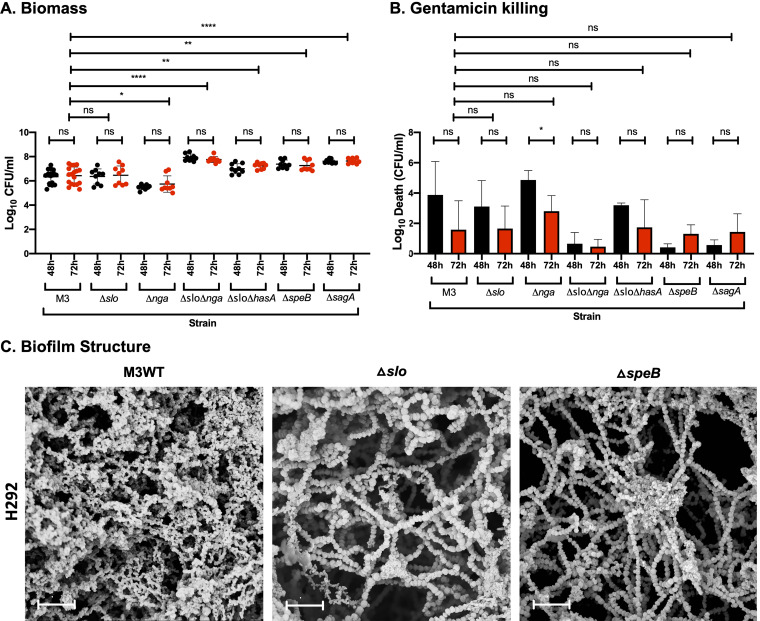
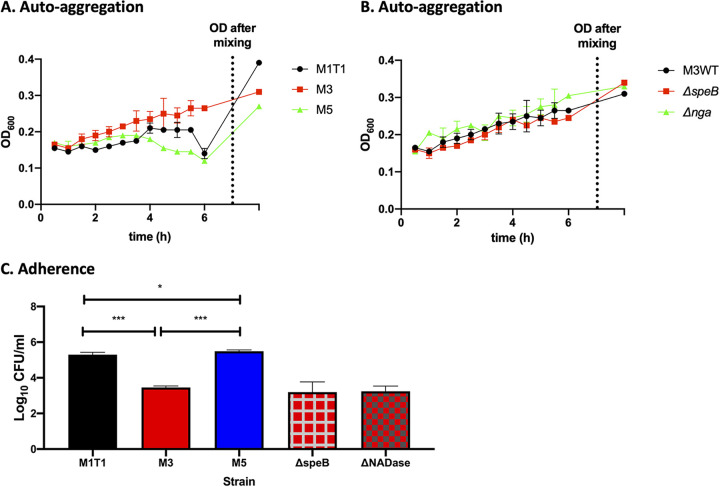
Biofilm formation by Streptococcus pyogenes (group A streptococcus [GAS]) in model systems mimicking the respiratory tract is poorly documented. Most studies have been conducted on abiotic surfaces, which poorly represent human tissues. We have previously shown that GAS forms mature and antibiotic-resistant biofilms on physiologically relevant epithelial cells. However, the roles of the substratum, extracellular matrix (ECM) components, and GAS virulence factors in biofilm formation and structure are unclear. In this study, biofilm formation was measured on respiratory epithelial cells and keratinocytes by determining biomass and antibiotic resistance, and biofilm morphology was visualized using scanning electron microscopy. All GAS isolates tested formed biofilms that had similar, albeit not identical, biomass and antibiotic resistance for both cell types. Interestingly, functionally mature biofilms formed more rapidly on keratinocytes but were structurally denser and coated with more ECM on respiratory epithelial cells. The ECM was crucial for biofilm integrity, as protein- and DNA-degrading enzymes induced bacterial release from biofilms. Abiotic surfaces supported biofilm formation, but these biofilms were structurally less dense and organized. No major role for M protein, capsule, or streptolysin O was observed in biofilm formation on epithelial cells, although some morphological differences were detected. NAD-glycohydrolase was required for optimal biofilm formation, whereas streptolysin S and cysteine protease SpeB impaired this process. Finally, no correlation was found between cell adherence or autoaggregation and GAS biofilm formation. Combined, these results provide a better understanding of the role of biofilm formation in GAS pathogenesis and can potentially provide novel targets for future treatments against GAS infections.
Keywords: Streptococcus pyogenes; adherence; aggregation; antibiotic resistance; biofilm formation; biofilm structure; biofilms; epithelial cells; extracellular matrix; keratinocytes; mucosal pathogens; respiratory tract; virulence factors.
Copyright © 2020 American Society for Microbiology.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
