

Defective endoplasmic reticulum-mitochondria contacts and bioenergetics in SEPN1-related myopathy
- PMID: 32661288
- PMCID: PMC7853070
- DOI: 10.1038/s41418-020-0587-z
Defective endoplasmic reticulum-mitochondria contacts and bioenergetics in SEPN1-related myopathy
Abstract
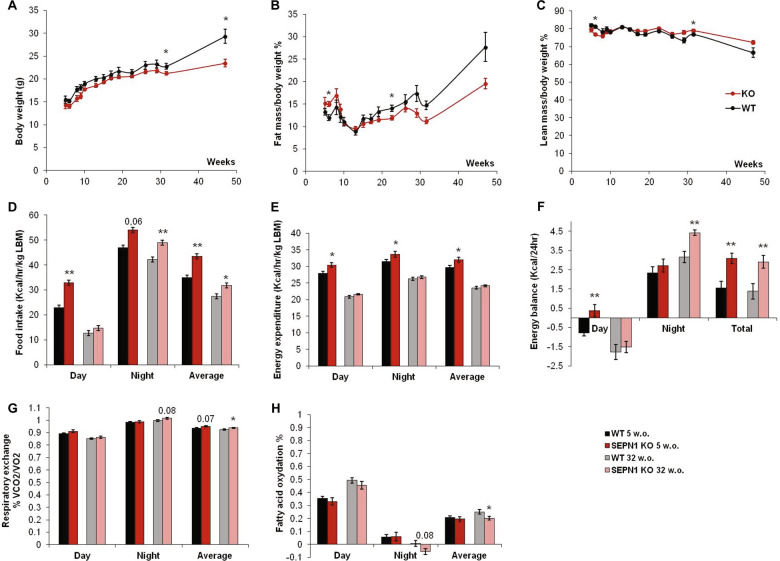
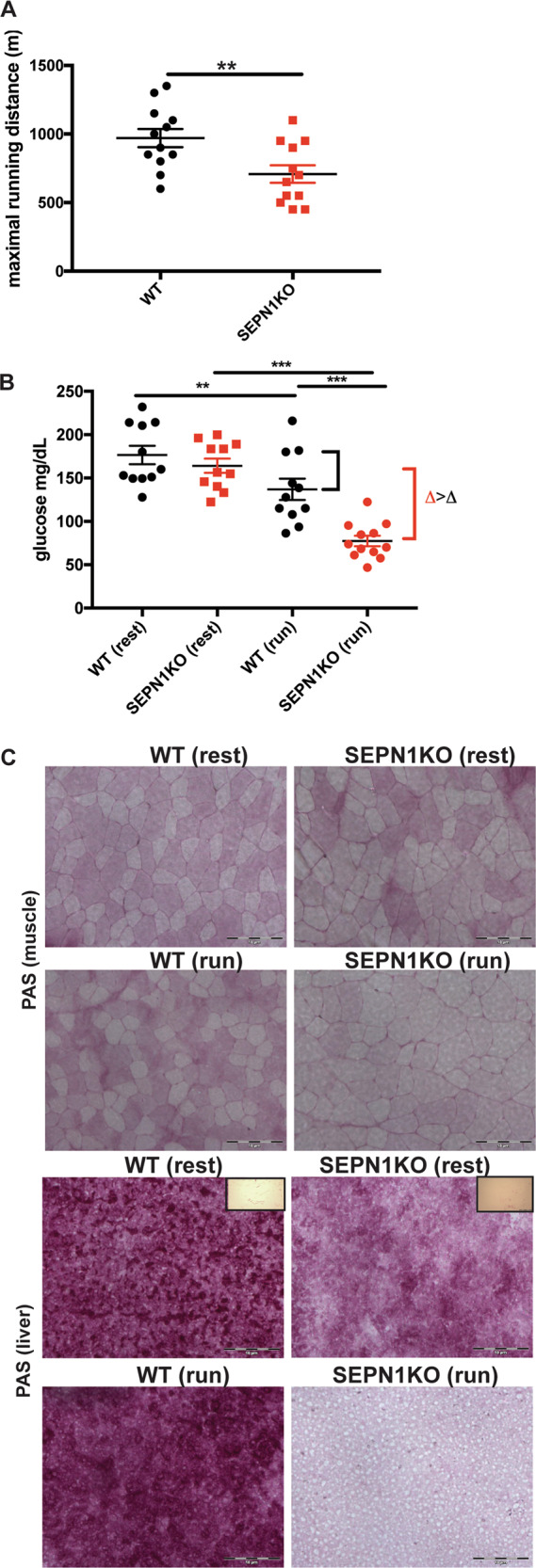
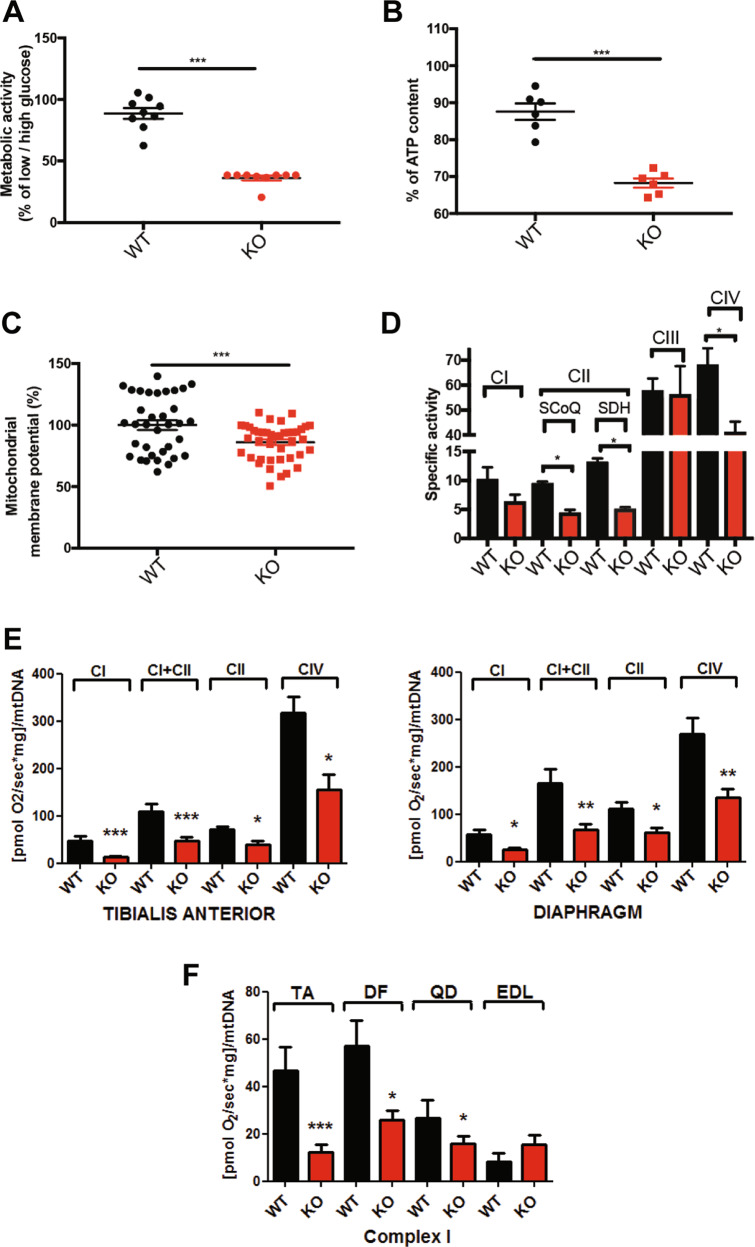
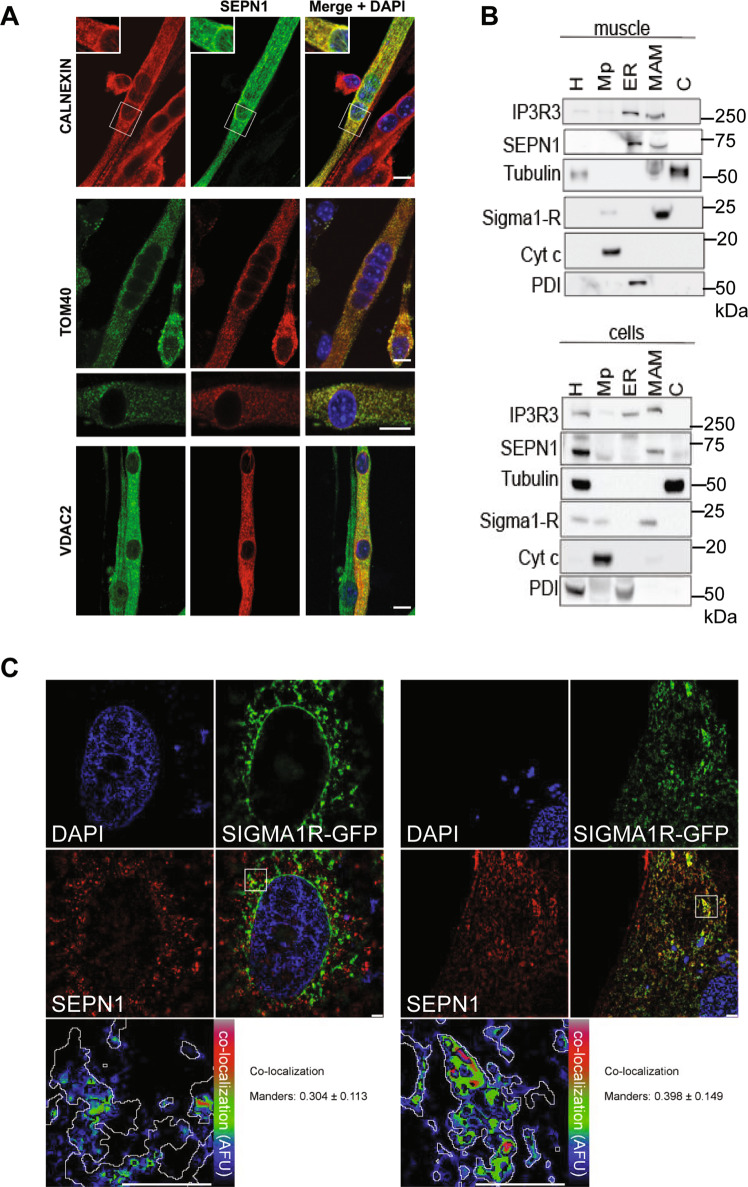
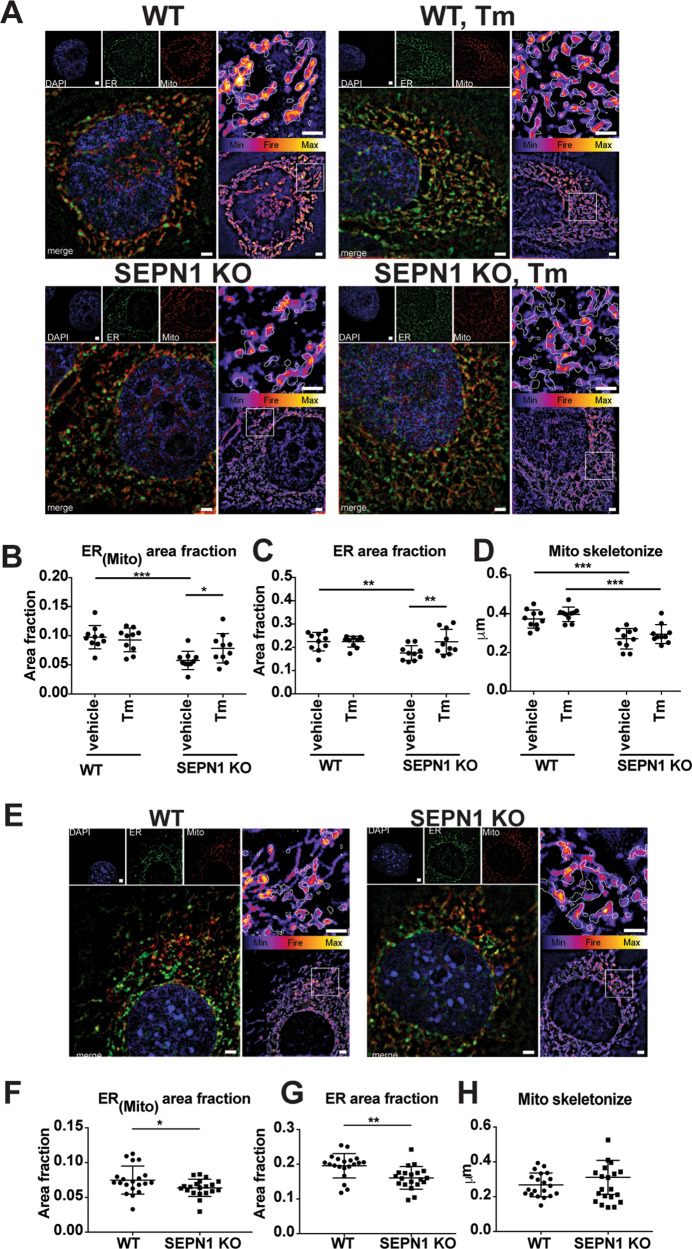
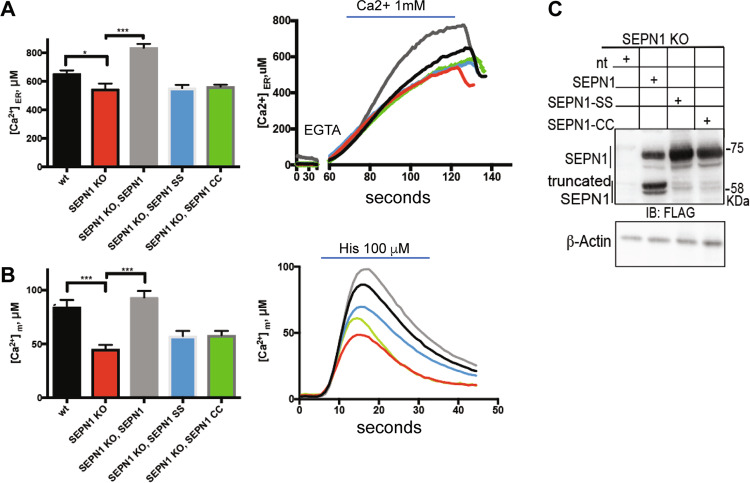
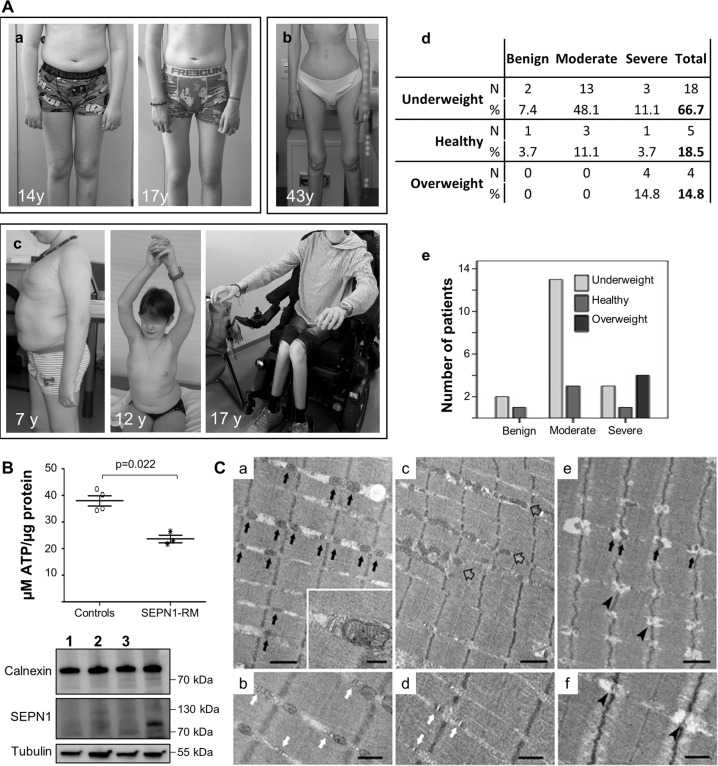
SEPN1-related myopathy (SEPN1-RM) is a muscle disorder due to mutations of the SEPN1 gene, which is characterized by muscle weakness and fatigue leading to scoliosis and life-threatening respiratory failure. Core lesions, focal areas of mitochondria depletion in skeletal muscle fibers, are the most common histopathological lesion. SEPN1-RM underlying mechanisms and the precise role of SEPN1 in muscle remained incompletely understood, hindering the development of biomarkers and therapies for this untreatable disease. To investigate the pathophysiological pathways in SEPN1-RM, we performed metabolic studies, calcium and ATP measurements, super-resolution and electron microscopy on in vivo and in vitro models of SEPN1 deficiency as well as muscle biopsies from SEPN1-RM patients. Mouse models of SEPN1 deficiency showed marked alterations in mitochondrial physiology and energy metabolism, suggesting that SEPN1 controls mitochondrial bioenergetics. Moreover, we found that SEPN1 was enriched at the mitochondria-associated membranes (MAM), and was needed for calcium transients between ER and mitochondria, as well as for the integrity of ER-mitochondria contacts. Consistently, loss of SEPN1 in patients was associated with alterations in body composition which correlated with the severity of muscle weakness, and with impaired ER-mitochondria contacts and low ATP levels. Our results indicate a role of SEPN1 as a novel MAM protein involved in mitochondrial bioenergetics. They also identify a systemic bioenergetic component in SEPN1-RM and establish mitochondria as a novel therapeutic target. This role of SEPN1 contributes to explain the fatigue and core lesions in skeletal muscle as well as the body composition abnormalities identified as part of the SEPN1-RM phenotype. Finally, these results point out to an unrecognized interplay between mitochondrial bioenergetics and ER homeostasis in skeletal muscle. They could therefore pave the way to the identification of biomarkers and therapeutic drugs for SEPN1-RM and for other disorders in which muscle ER-mitochondria cross-talk are impaired.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
