

RAD51 Gene Family Structure and Function
- PMID: 32663049
- PMCID: PMC7703940
- DOI: 10.1146/annurev-genet-021920-092410
RAD51 Gene Family Structure and Function
Abstract
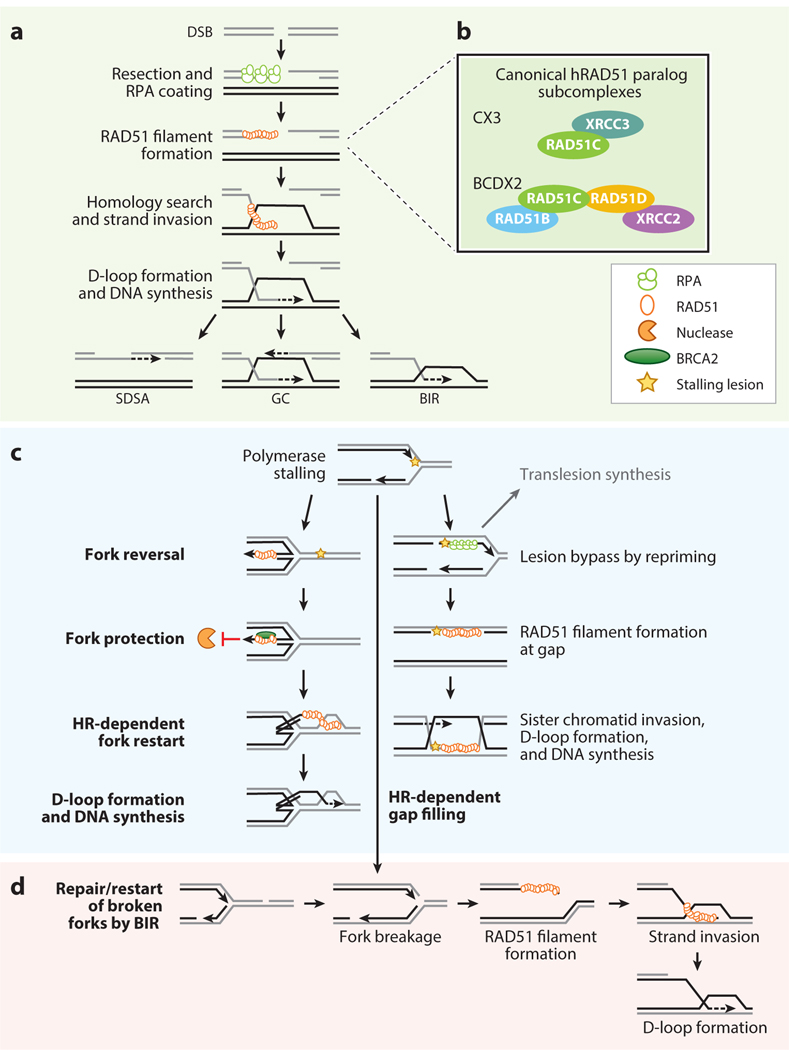
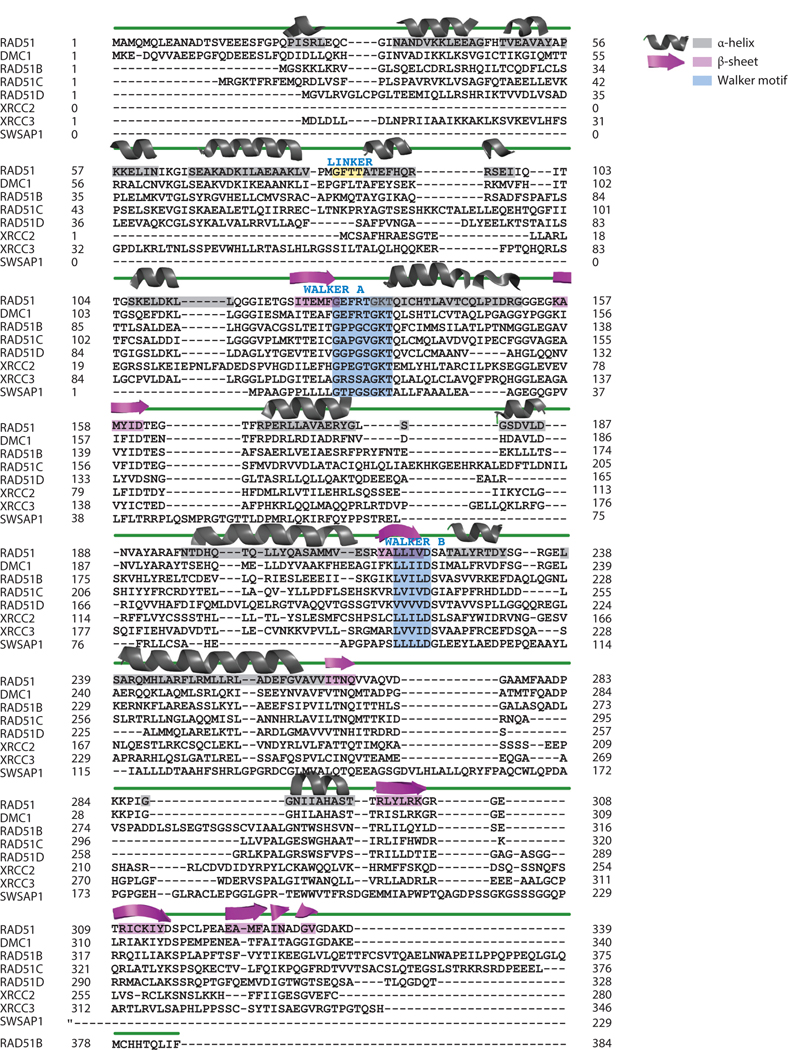
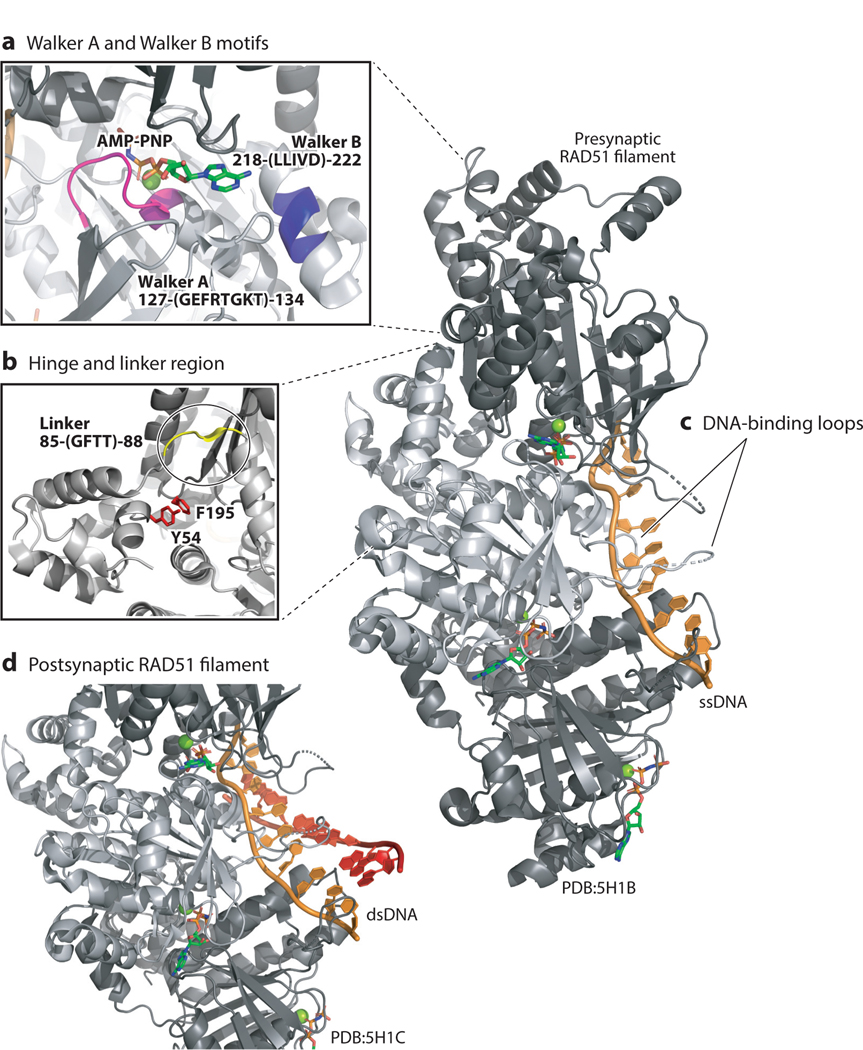
Accurate DNA repair and replication are critical for genomic stability and cancer prevention. RAD51 and its gene family are key regulators of DNA fidelity through diverse roles in double-strand break repair, replication stress, and meiosis. RAD51 is an ATPase that forms a nucleoprotein filament on single-stranded DNA. RAD51 has the function of finding and invading homologous DNA sequences to enable accurate and timely DNA repair. Its paralogs, which arose from ancient gene duplications of RAD51, have evolved to regulate and promote RAD51 function. Underscoring its importance, misregulation of RAD51, and its paralogs, is associated with diseases such as cancer and Fanconi anemia. In this review, we focus on the mammalian RAD51 structure and function and highlight the use of model systems to enable mechanistic understanding of RAD51 cellular roles. We also discuss how misregulation of the RAD51 gene family members contributes to disease and consider new approaches to pharmacologically inhibit RAD51.
Keywords: RAD51; RAD51 paralog; Shu complex; double-strand break repair; homologous recombination; replication.
Figures
References
-
- Aten JA, Stap J, Krawczyk PM, van Oven CH, Hoebe RA, et al. 2004. Dynamics of DNA double-strand breaks revealed by clustering of damaged chromosome domains. Science 303:92–95 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
