

Composition, structure and function of the corneal stroma
- PMID: 32663498
- PMCID: PMC7508887
- DOI: 10.1016/j.exer.2020.108137
Composition, structure and function of the corneal stroma
Abstract
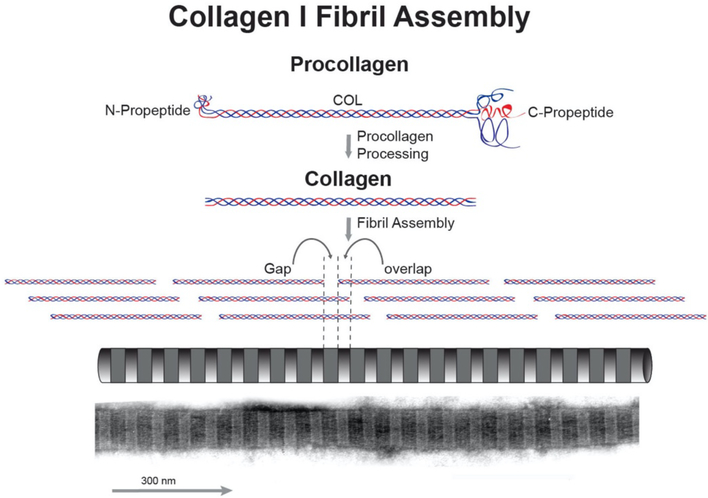
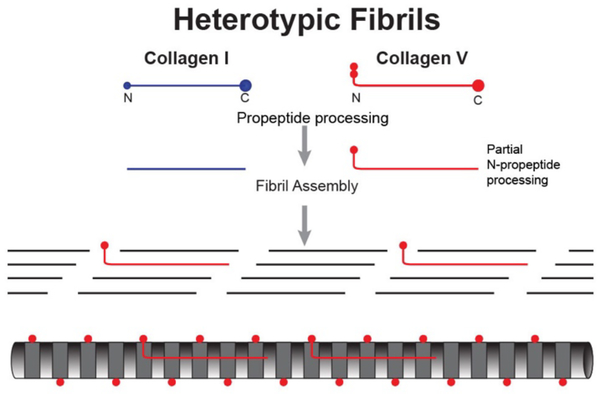
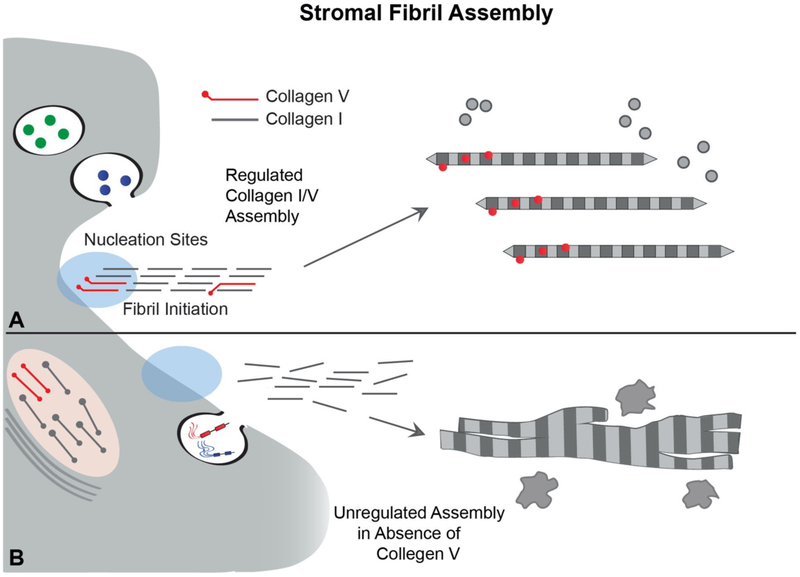
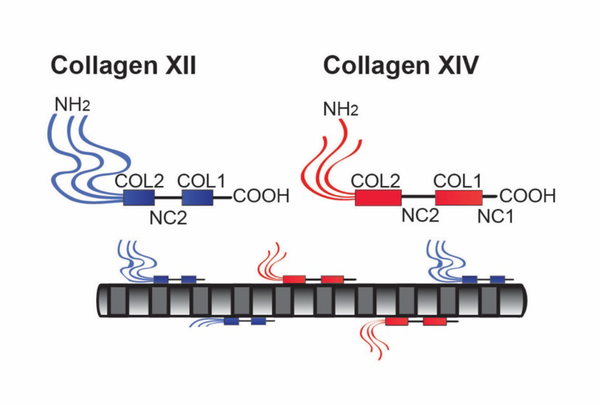
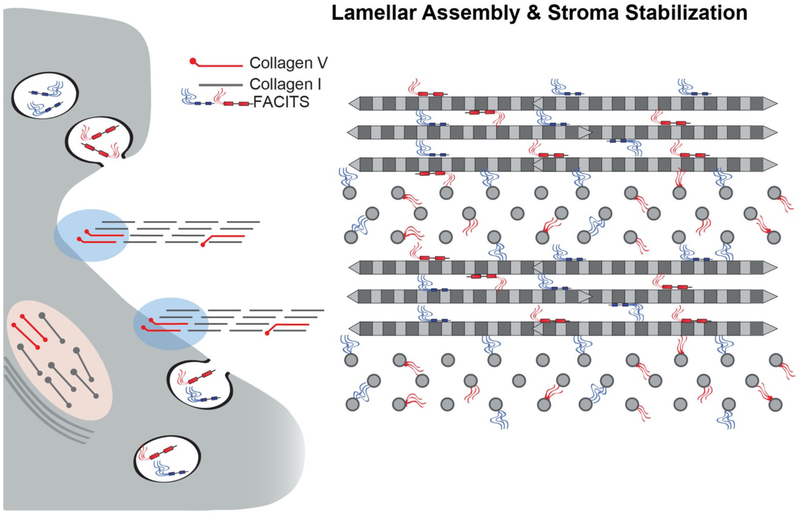
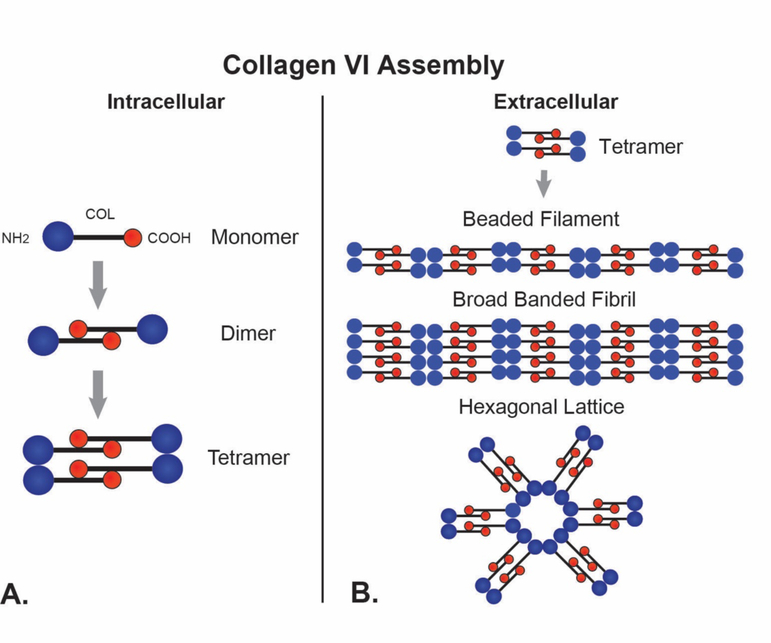
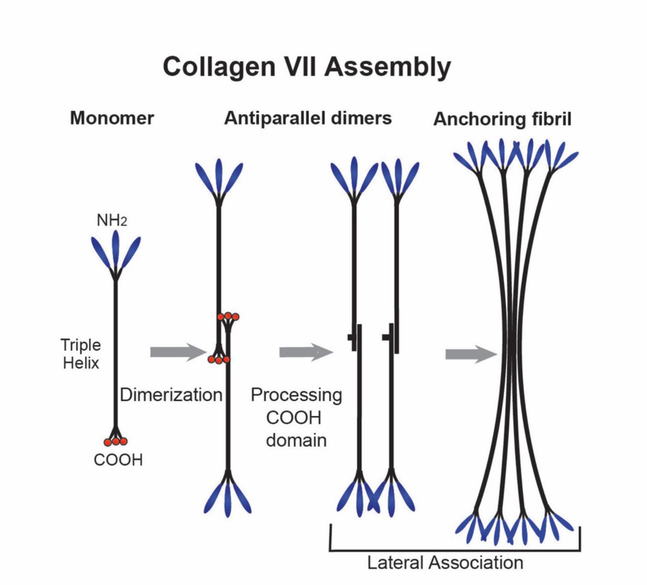
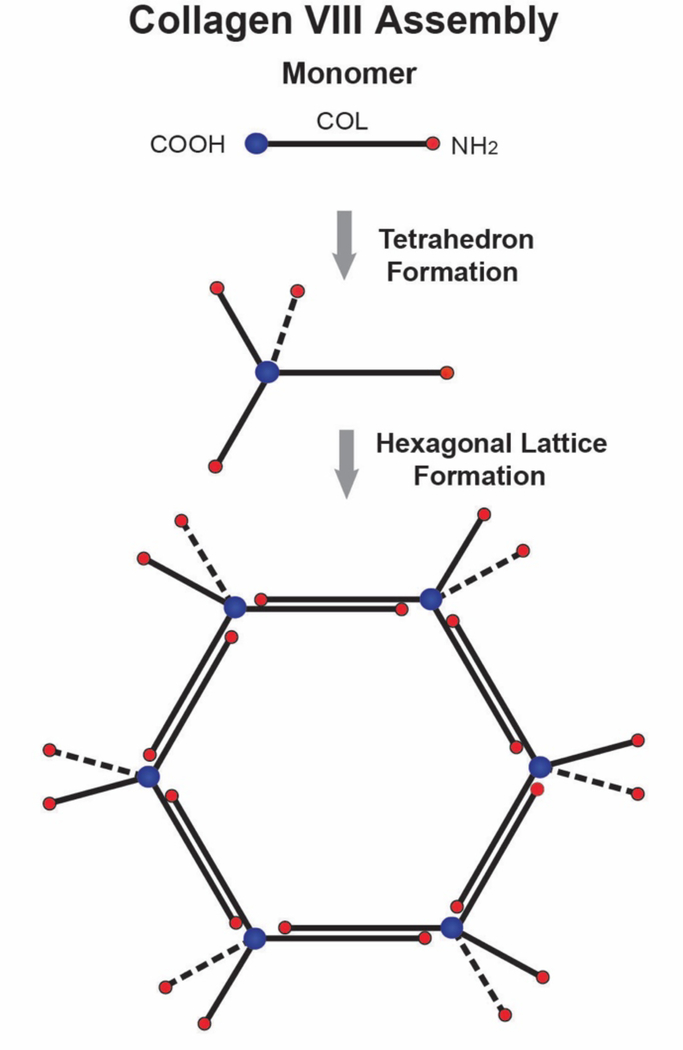
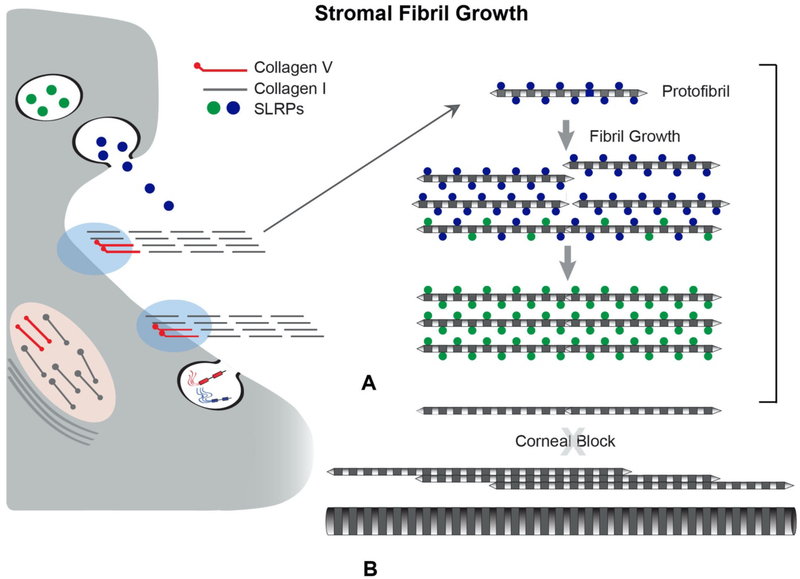
No other tissue in the body depends more on the composition and organization of the extracellular matrix (ECM) for normal structure and function than the corneal stroma. The precise arrangement and orientation of collagen fibrils, lamellae and keratocytes that occurs during development and is needed in adults to maintain stromal function is dependent on the regulated interaction of multiple ECM components that contribute to attain the unique properties of the cornea: transparency, shape, mechanical strength, and avascularity. This review summarizes the contribution of different ECM components, their structure, regulation and function in modulating the properties of the corneal stroma. Fibril forming collagens (I, III, V), fibril associated collagens with interrupted triple helices (XII and XIV), network forming collagens (IV, VI and VIII) as well as small leucine-rich proteoglycans (SLRP) expressed in the stroma: decorin, biglycan, lumican, keratocan, and fibromodulin are some of the ECM components reviewed in this manuscript. There are spatial and temporal differences in the expression of these ECM components, as well as interactions among them that contribute to stromal function. Unique regions within the stroma like Bowman's layer and Descemet's layer are discussed. To define the complexity of corneal stroma composition and structure as well as the relationship to function is a daunting task. Our knowledge is expanding, and we expect that this review provides a comprehensive overview of current knowledge, definition of gaps and suggests future research directions.
Keywords: Collagen fibril; Collagens; Composition; Cornea; Proteoglycans; Stroma; Structure.
Copyright © 2020 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures
References
-
- Akama TO, Nishida K, Nakayama J, Watanabe H, Ozaki K, Nakamura T, Dota A, Kawasaki S, Inoue Y, Maeda N, Yamamoto S, Fujiwara T, Thonar EJ, Shimomura Y, Kinoshita S, Tanigami A, Fukuda MN, 2000. Macular corneal dystrophy type I and type II are caused by distinct mutations in a new sulphotransferase gene. Nat Genet 26, 237–241. - PubMed
-
- Arai K, Nagashima Y, Takemoto T, Nishiyama T, 2008. Mechanical strain increases expression of type XII collagen in murine osteoblastic MC3T3-E1 cells. Cell Struct Funct 33, 203–210. - PubMed
-
- Armstrong DJ, Hiscott P, Batterbury M, Kaye S, 2003. Keratocyte matrix interactions and thrombospondin 2. Mol Vis 9, 74–79. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
