

Conditional DnaB Protein Splicing Is Reversibly Inhibited by Zinc in Mycobacteria
- PMID: 32665276
- PMCID: PMC7360933
- DOI: 10.1128/mBio.01403-20
Conditional DnaB Protein Splicing Is Reversibly Inhibited by Zinc in Mycobacteria
Abstract
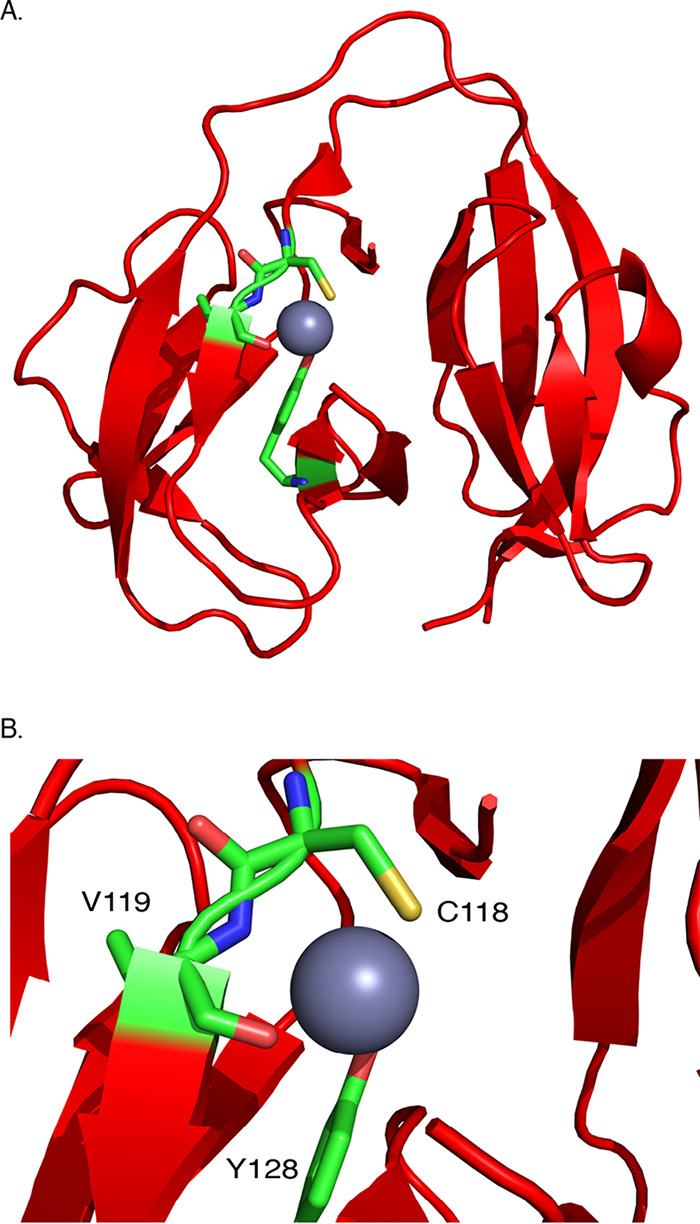
Inteins, as posttranslational regulatory elements, can tune protein function to environmental changes by conditional protein splicing (CPS). Translated as subdomains interrupting host proteins, inteins splice to scarlessly join flanking sequences (exteins). We used DnaB-intein1 (DnaBi1) from a replicative helicase of Mycobacterium smegmatis to build a
Keywords: DNA helicase; conditional protein splicing; intein; mycobacteria.
Copyright © 2020 Woods et al.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
