

The mevalonate pathway is an actionable vulnerability of t(4;14)-positive multiple myeloma
- PMID: 32665698
- PMCID: PMC7359767
- DOI: 10.1038/s41375-020-0962-2
The mevalonate pathway is an actionable vulnerability of t(4;14)-positive multiple myeloma
Abstract
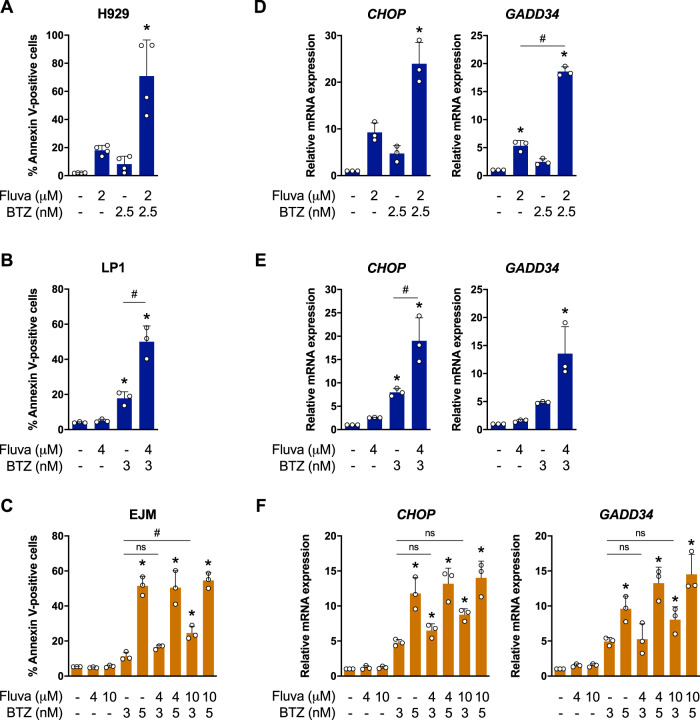
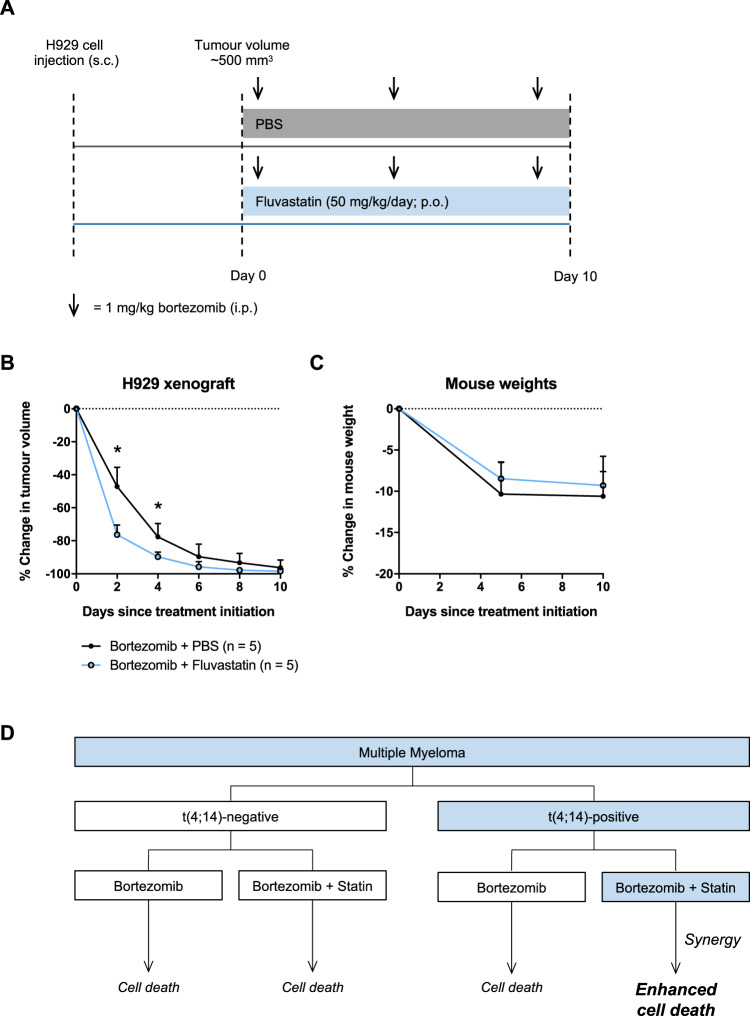
Multiple myeloma (MM) is a plasma cell malignancy that is often driven by chromosomal translocations. In particular, patients with t(4;14)-positive disease have worse prognosis compared to other MM subtypes. Herein, we demonstrated that t(4;14)-positive cells are highly dependent on the mevalonate (MVA) pathway for survival. Moreover, we showed that this metabolic vulnerability is immediately actionable, as inhibiting the MVA pathway with a statin preferentially induced apoptosis in t(4;14)-positive cells. In response to statin treatment, t(4;14)-positive cells activated the integrated stress response (ISR), which was augmented by co-treatment with bortezomib, a proteasome inhibitor. We identified that t(4;14)-positive cells depend on the MVA pathway for the synthesis of geranylgeranyl pyrophosphate (GGPP), as exogenous GGPP fully rescued statin-induced ISR activation and apoptosis. Inhibiting protein geranylgeranylation similarly induced the ISR in t(4;14)-positive cells, suggesting that this subtype of MM depends on GGPP, at least in part, for protein geranylgeranylation. Notably, fluvastatin treatment synergized with bortezomib to induce apoptosis in t(4;14)-positive cells and potentiated the anti-tumor activity of bortezomib in vivo. Our data implicate the t(4;14) translocation as a biomarker of statin sensitivity and warrant further clinical evaluation of a statin in combination with bortezomib for the treatment of t(4;14)-positive disease.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
