

Lipid kinases PIP5K7 and PIP5K9 are required for polyamine-triggered K+ efflux in Arabidopsis roots
- PMID: 32666545
- PMCID: PMC7693229
- DOI: 10.1111/tpj.14932
Lipid kinases PIP5K7 and PIP5K9 are required for polyamine-triggered K+ efflux in Arabidopsis roots
Abstract
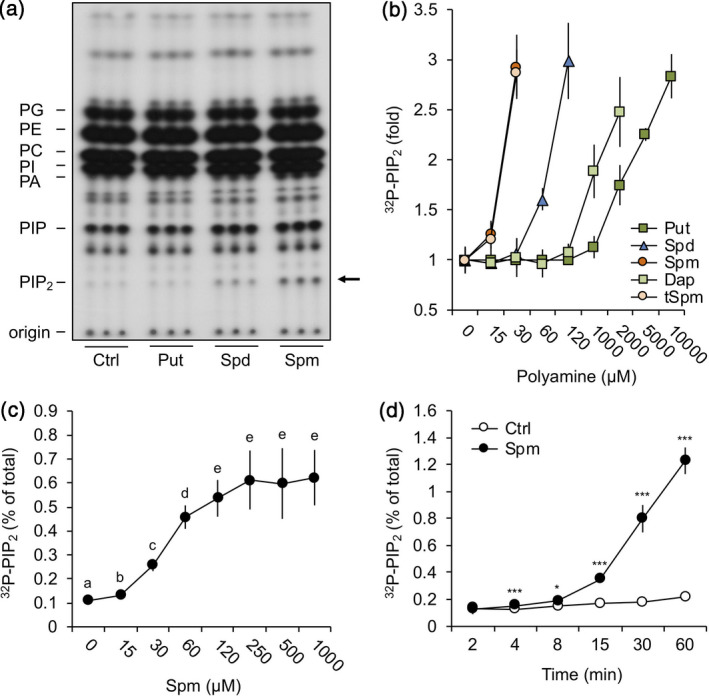
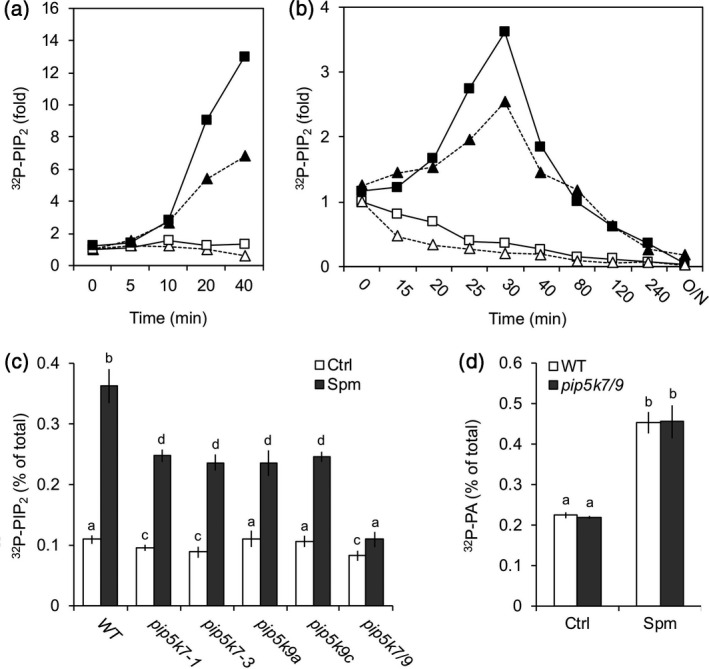
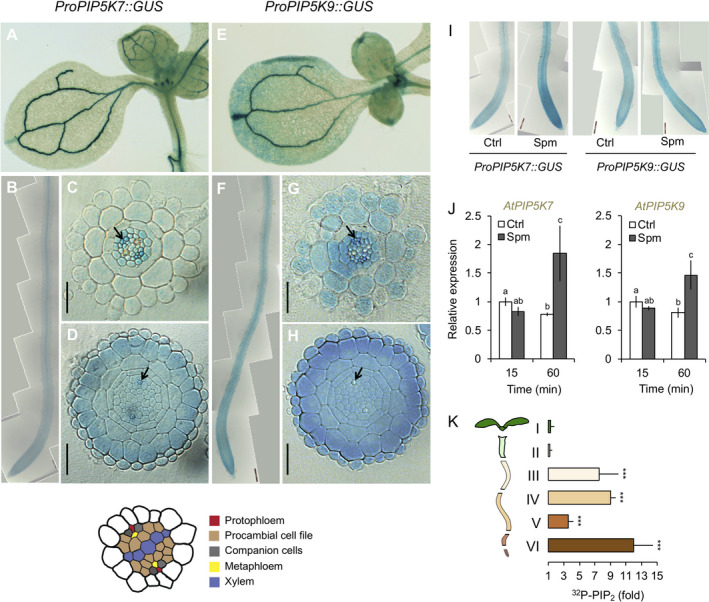
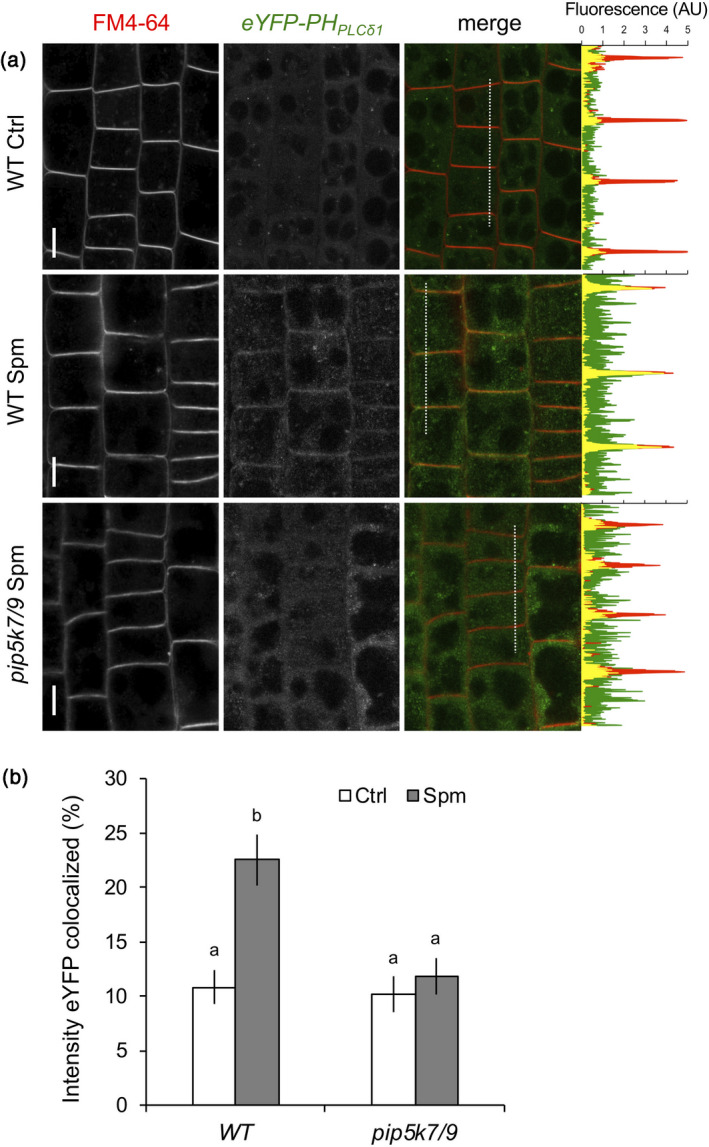
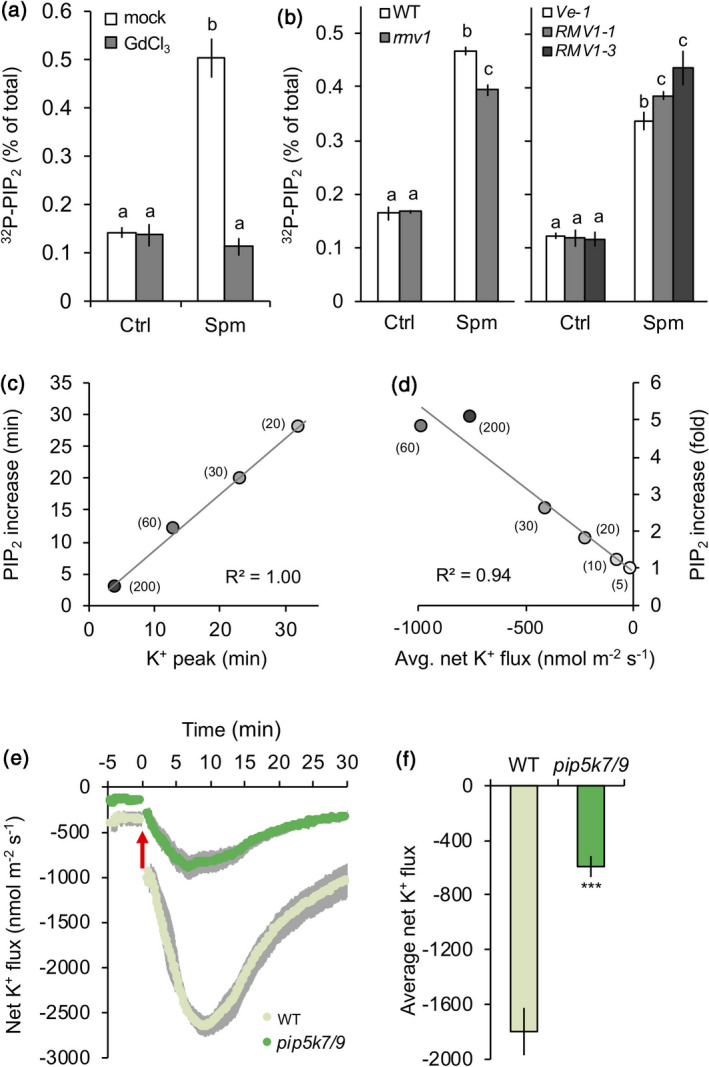
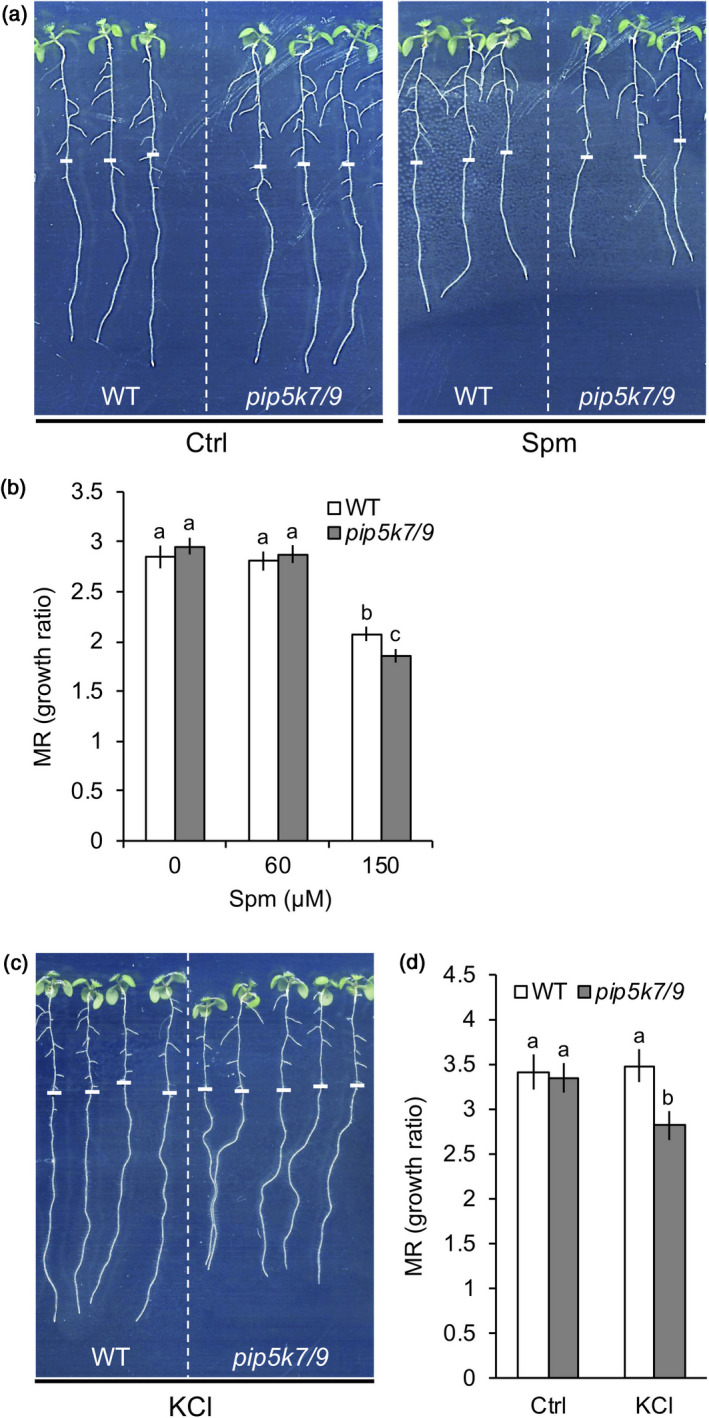
Polyamines, such as putrescine, spermidine and spermine (Spm), are low-molecular-weight polycationic molecules present in all living organisms. Despite their implication in plant cellular processes, little is known about their molecular mode of action. Here, we demonstrate that polyamines trigger a rapid increase in the regulatory membrane lipid phosphatidylinositol 4,5-bisphosphate (PIP2 ), and that this increase is required for polyamine effects on K+ efflux in Arabidopsis roots. Using in vivo 32 Pi -labelling of Arabidopsis seedlings, low physiological (μm) concentrations of Spm were found to promote a rapid PIP2 increase in roots that was time- and dose-dependent. Confocal imaging of a genetically encoded PIP2 biosensor revealed that this increase was triggered at the plasma membrane. Differential 32 Pi -labelling suggested that the increase in PIP2 was generated through activation of phosphatidylinositol 4-phosphate 5-kinase (PIP5K) activity rather than inhibition of a phospholipase C or PIP2 5-phosphatase activity. Systematic analysis of transfer DNA insertion mutants identified PIP5K7 and PIP5K9 as the main candidates involved in the Spm-induced PIP2 response. Using non-invasive microelectrode ion flux estimation, we discovered that the Spm-triggered K+ efflux response was strongly reduced in pip5k7 pip5k9 seedlings. Together, our results provide biochemical and genetic evidence for a physiological role of PIP2 in polyamine-mediated signalling controlling K+ flux in plants.
Keywords: Arabidopsis; K+ flux; phosphatidic acid (PA); phosphatidylinositol 4,5-bisphosphate (PIP2); phosphatidylinositol 4-phosphate 5-kinase (PIP5K); phosphoinositide signalling; phospholipids; polyamines.
© 2020 The Authors. The Plant Journal published by Society for Experimental Biology and John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare that they have no conflict of interests.
Figures
References
-
- Angelini, R. , Cona, A. , Federico, R. , Fincato, P. , Tavladoraki, P. and Tisi, A. (2010) Plant amine oxidases “on the move”: An update. Plant Physiol. Biochem. 48, 560–564. - PubMed
-
- Anschütz, U. , Becker, D. and Shabala, S. (2014) Going beyond nutrition: Regulation of potassium homeostasis as a common denominator of plant adaptive responses to environment. J. Plant Physiol. 171, 670–687. - PubMed
-
- Arisz, S.A. and Munnik, T. (2013) Distinguishing phosphatidic acid pools from de novo synthesis, PLD, and DGK. Methods Mol. Biol. 1009, 55–62. - PubMed
-
- Armengaud, P. , Zambaux, K. , Hills, A. , Sulpice, R. , Pattison, R.J. , Blatt, M.R. and Amtmann, A. (2009) EZ‐Rhizo: integrated software for the fast and accurate measurement of root system architecture. Plant J. 57, 945–956. - PubMed
-
- Athwal, G.S. and Huber, S.C. (2002) Divalent cations and polyamines bind to loop 8 of 14‐3‐3 proteins, modulating their interaction with phosphorylated nitrate reductase. Plant J. 29, 119–129. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
