

PARP1 catalytic variants reveal branching and chain length-specific functions of poly(ADP-ribose) in cellular physiology and stress response
- PMID: 32667640
- PMCID: PMC7544232
- DOI: 10.1093/nar/gkaa590
PARP1 catalytic variants reveal branching and chain length-specific functions of poly(ADP-ribose) in cellular physiology and stress response
Abstract
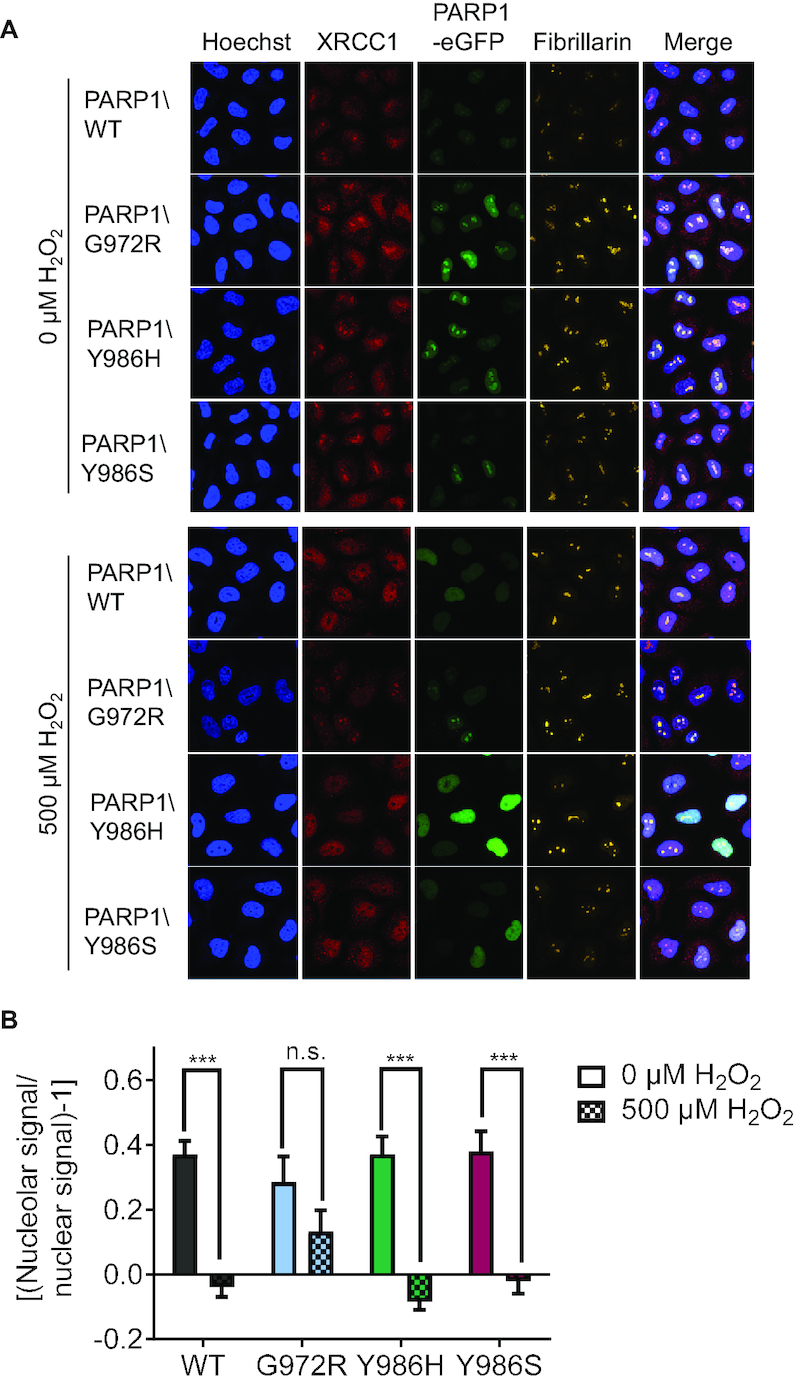
Poly(ADP-ribosyl)ation regulates numerous cellular processes like genome maintenance and cell death, thus providing protective functions but also contributing to several pathological conditions. Poly(ADP-ribose) (PAR) molecules exhibit a remarkable heterogeneity in chain lengths and branching frequencies, but the biological significance of this is basically unknown. To unravel structure-specific functions of PAR, we used PARP1 mutants producing PAR of different qualities, i.e. short and hypobranched (PARP1\G972R), short and moderately hyperbranched (PARP1\Y986S), or strongly hyperbranched PAR (PARP1\Y986H). By reconstituting HeLa PARP1 knockout cells, we demonstrate that PARP1\G972R negatively affects cellular endpoints, such as viability, cell cycle progression and genotoxic stress resistance. In contrast, PARP1\Y986S elicits only mild effects, suggesting that PAR branching compensates for short polymer length. Interestingly, PARP1\Y986H exhibits moderate beneficial effects on cell physiology. Furthermore, different PARP1 mutants have distinct effects on molecular processes, such as gene expression and protein localization dynamics of PARP1 itself, and of its downstream factor XRCC1. Finally, the biological relevance of PAR branching is emphasized by the fact that branching frequencies vary considerably during different phases of the DNA damage-induced PARylation reaction and between different mouse tissues. Taken together, this study reveals that PAR branching and chain length essentially affect cellular functions, which further supports the notion of a 'PAR code'.
© The Author(s) 2020. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures










References
-
- Leung A.K.L. PARPs. Curr. Biol. 2017; 27:R1256–R1258. - PubMed
-
- Luscher B., Butepage M., Eckei L., Krieg S., Verheugd P., Shilton B.H.. ADP-Ribosylation, a multifaceted posttranslational modification involved in the control of cell physiology in health and disease. Chem. Rev. 2018; 118:1092–1136. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
