

Temperature-dependent growth contributes to long-term cold sensing
- PMID: 32669706
- PMCID: PMC7116785
- DOI: 10.1038/s41586-020-2485-4
Temperature-dependent growth contributes to long-term cold sensing
Erratum in
-
Publisher Correction: Temperature-dependent growth contributes to long-term cold sensing.Nature. 2020 Sep;585(7824):E8. doi: 10.1038/s41586-020-2694-x. Nature. 2020. PMID: 32839616
Abstract
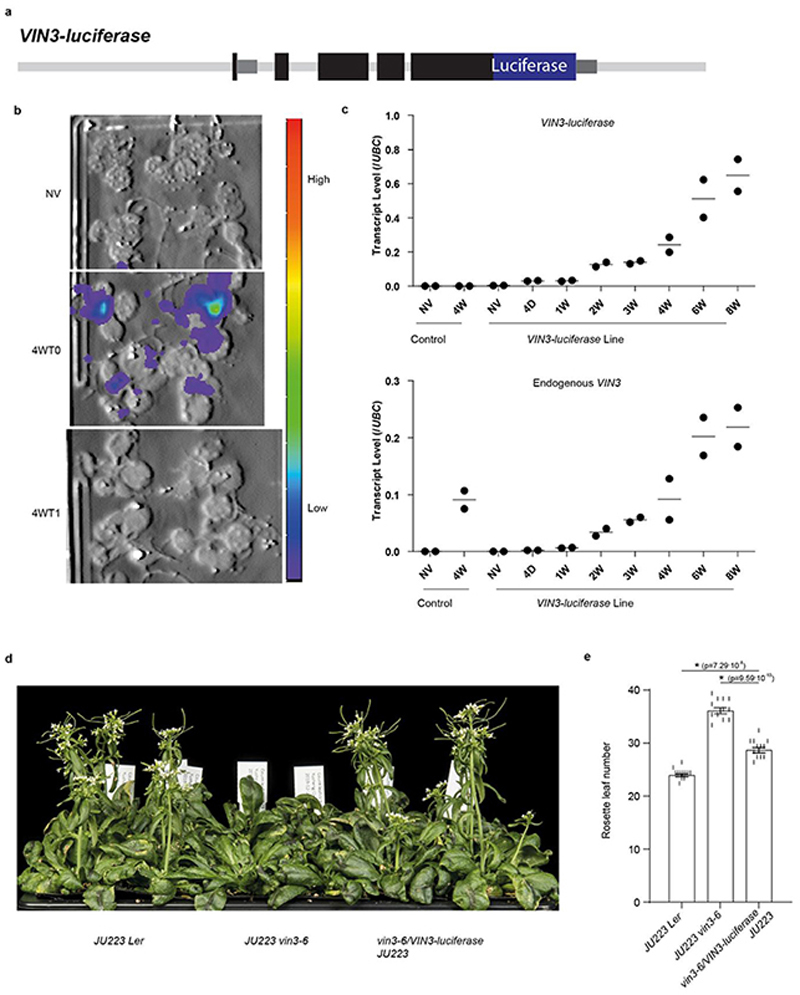
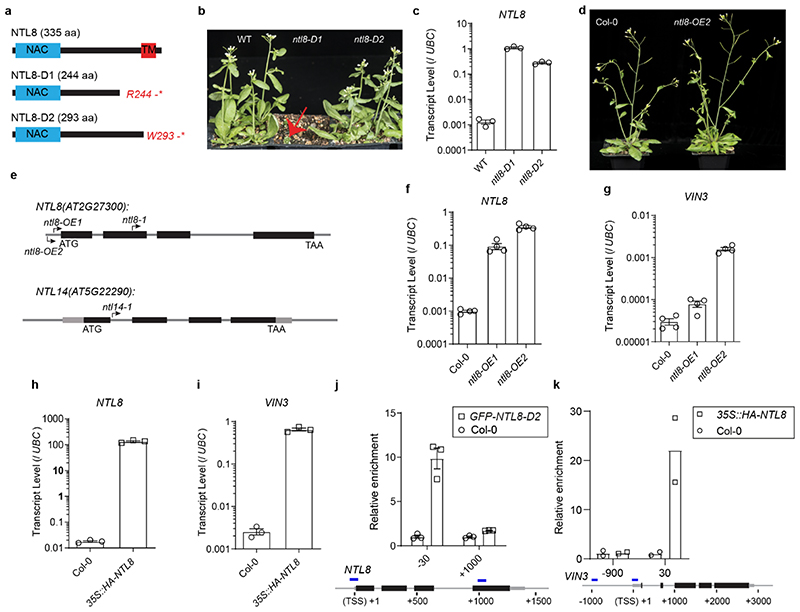
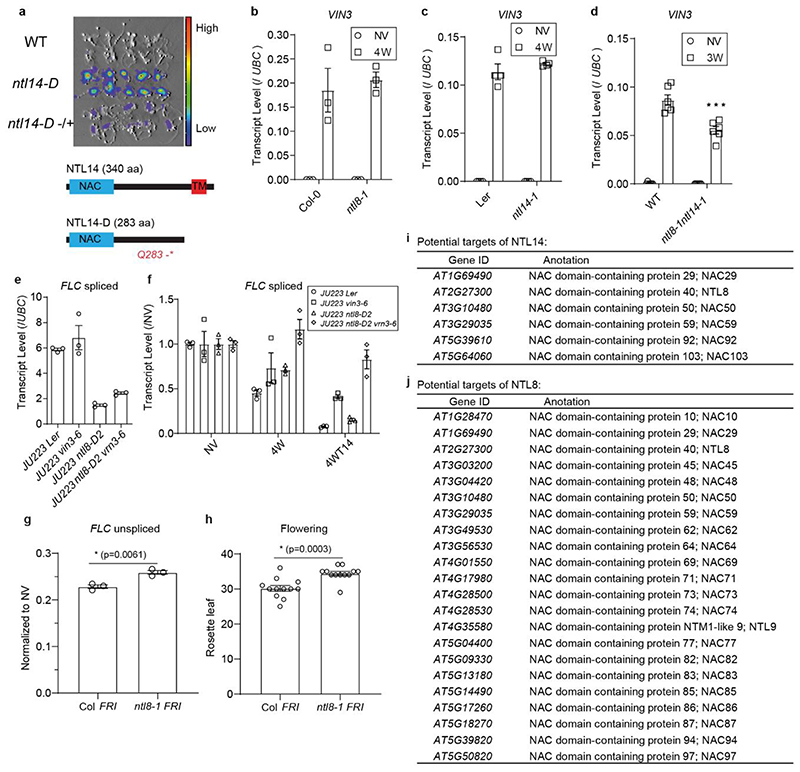
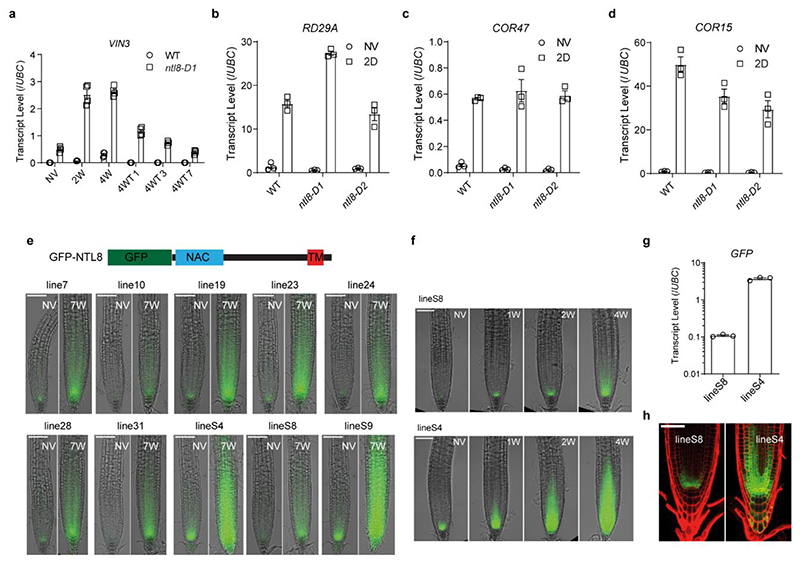
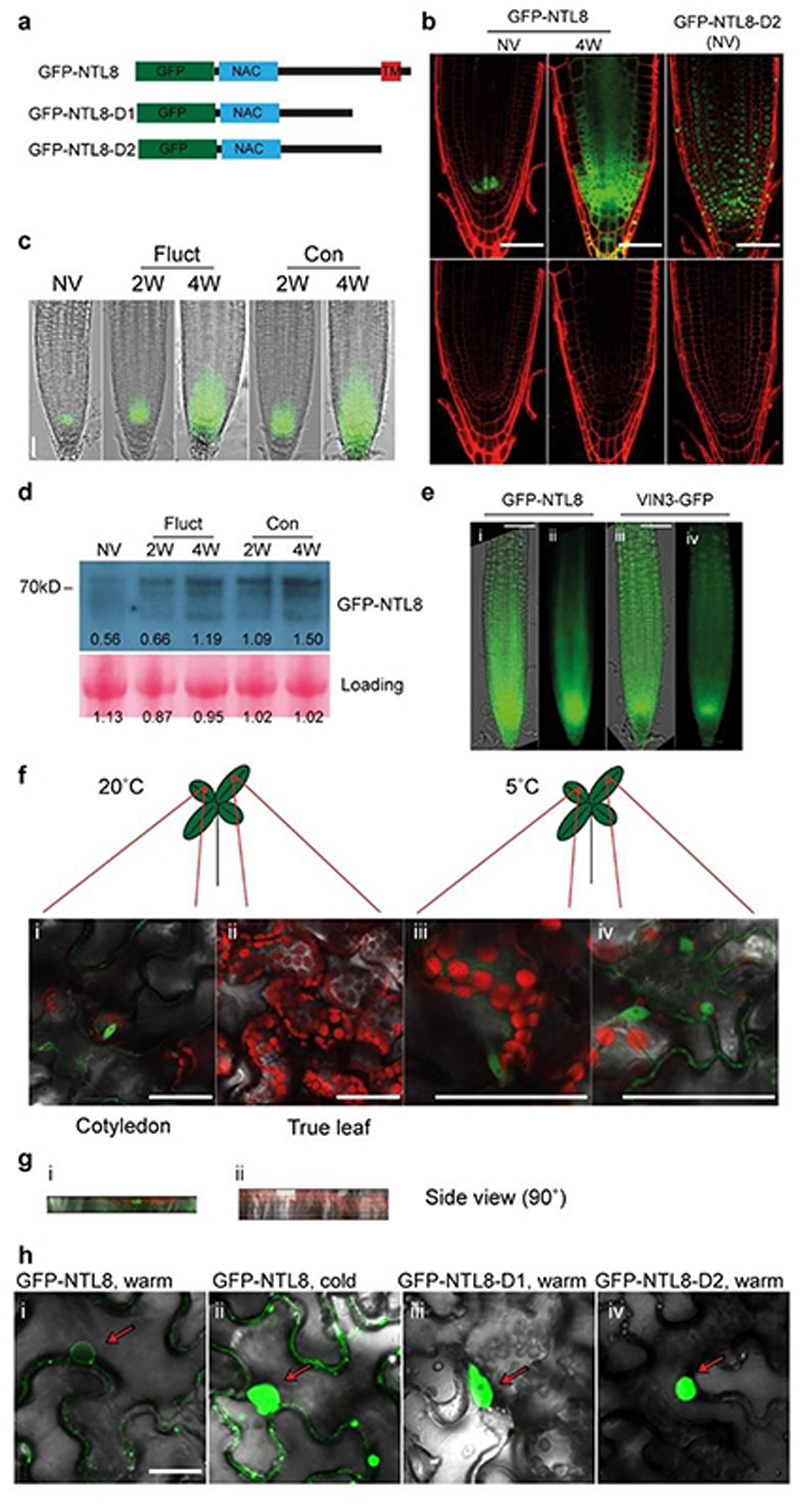
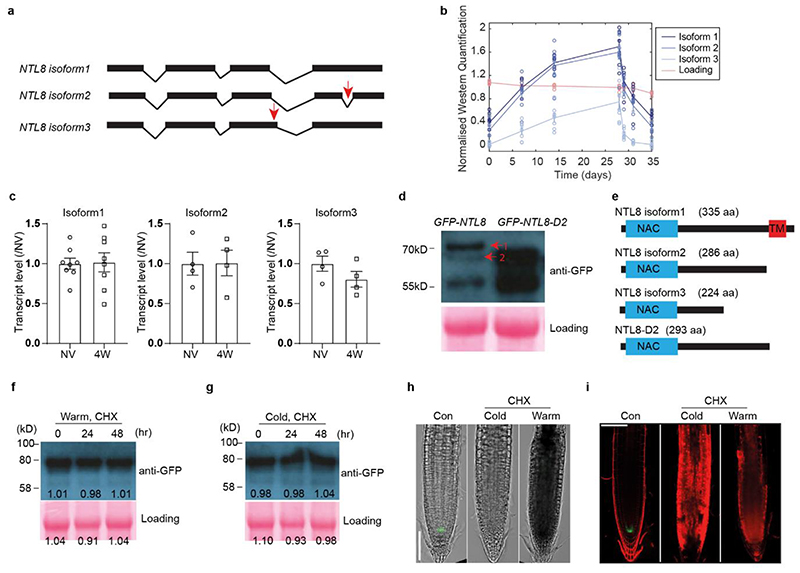
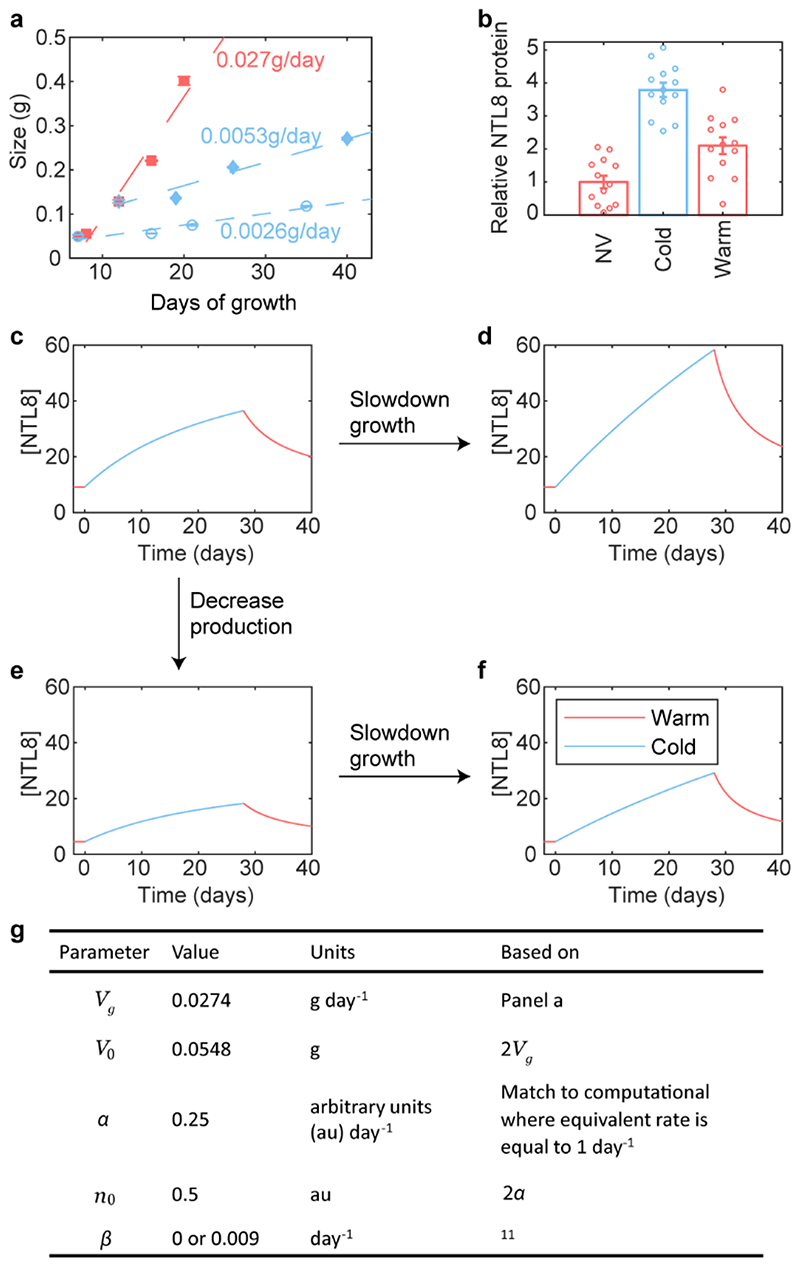
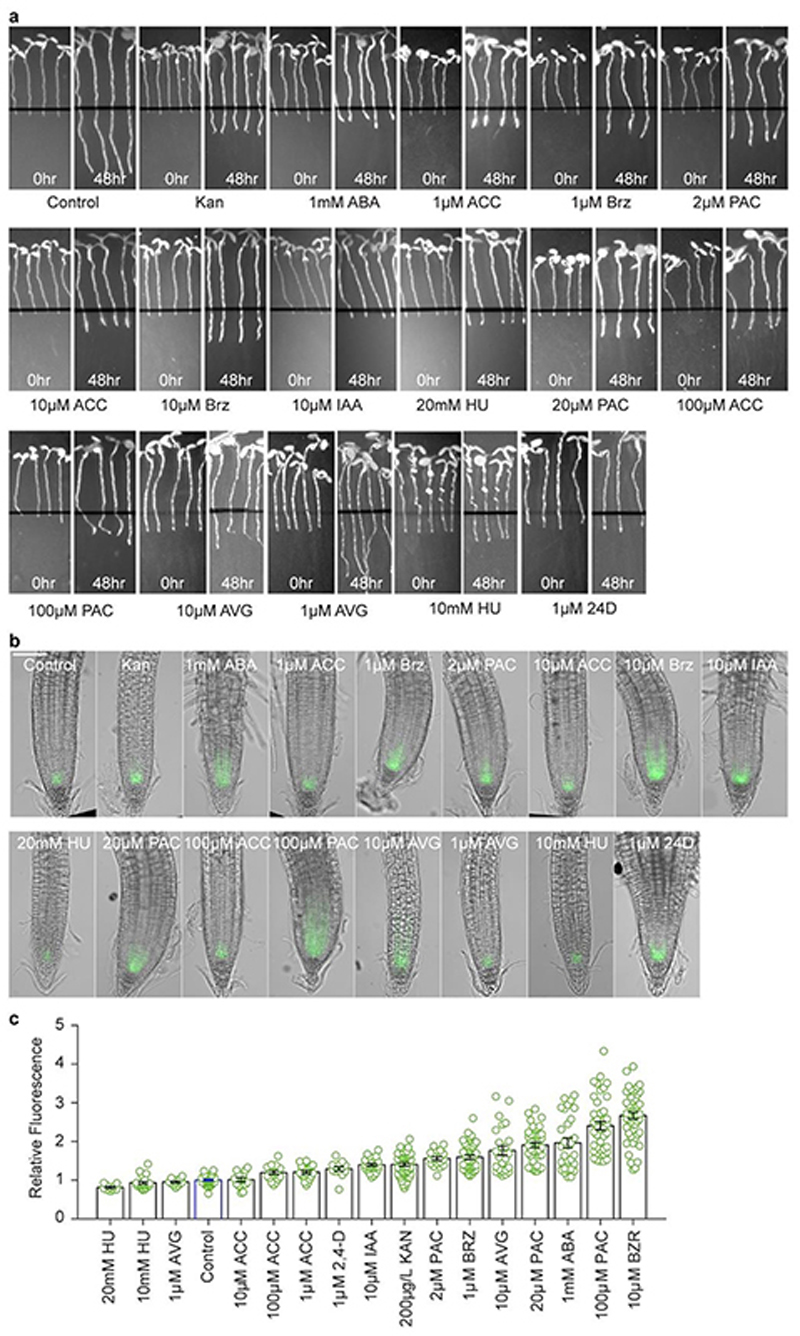
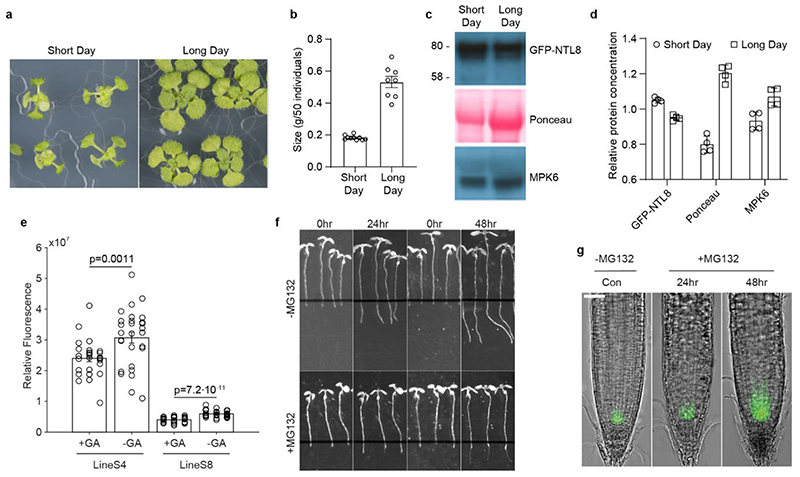
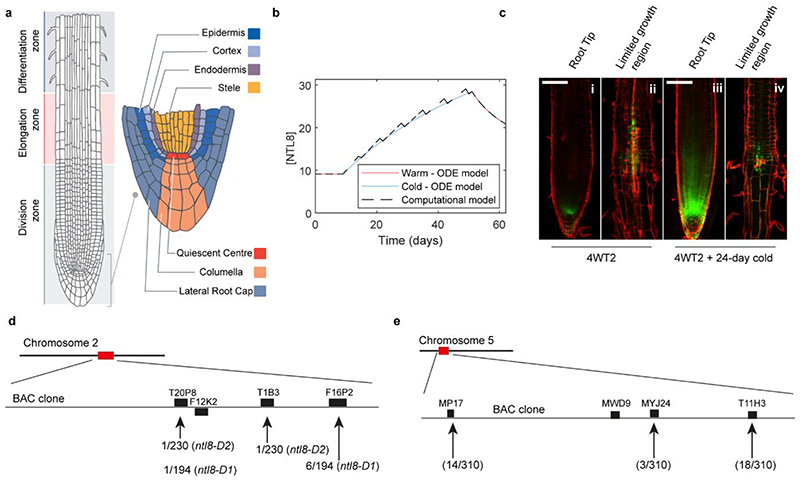
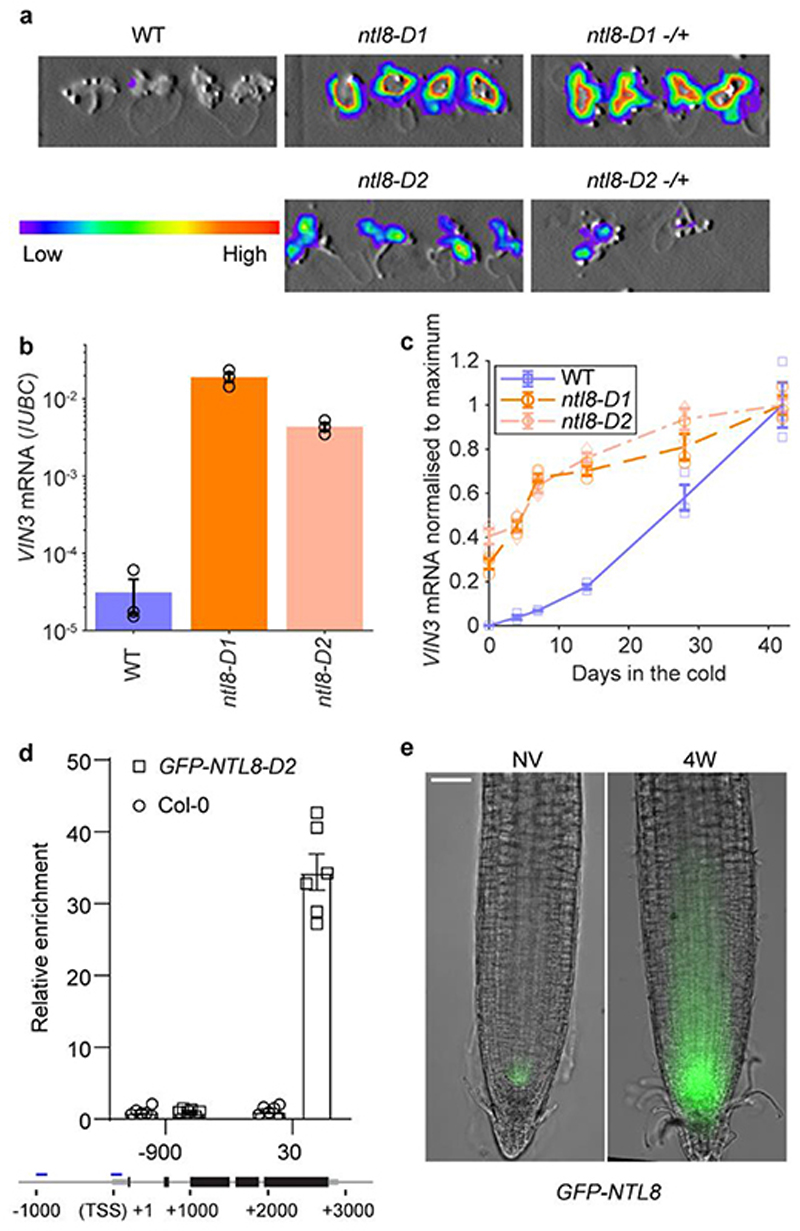
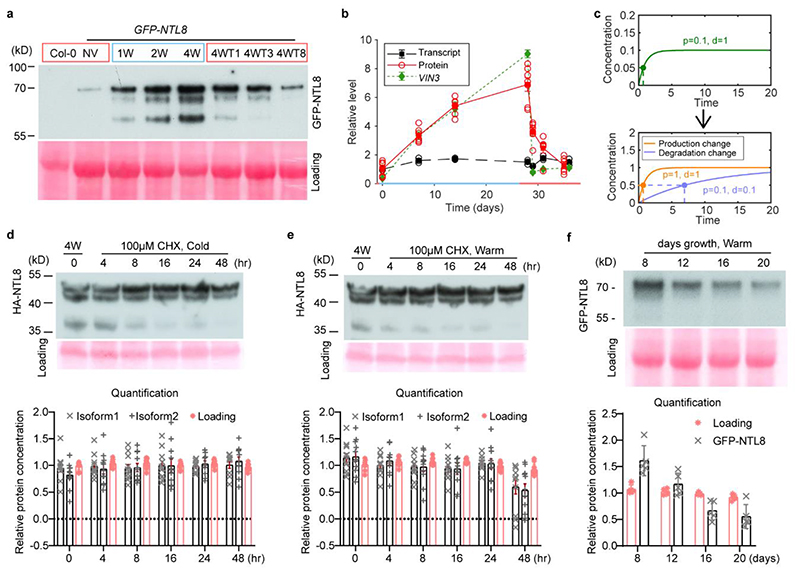
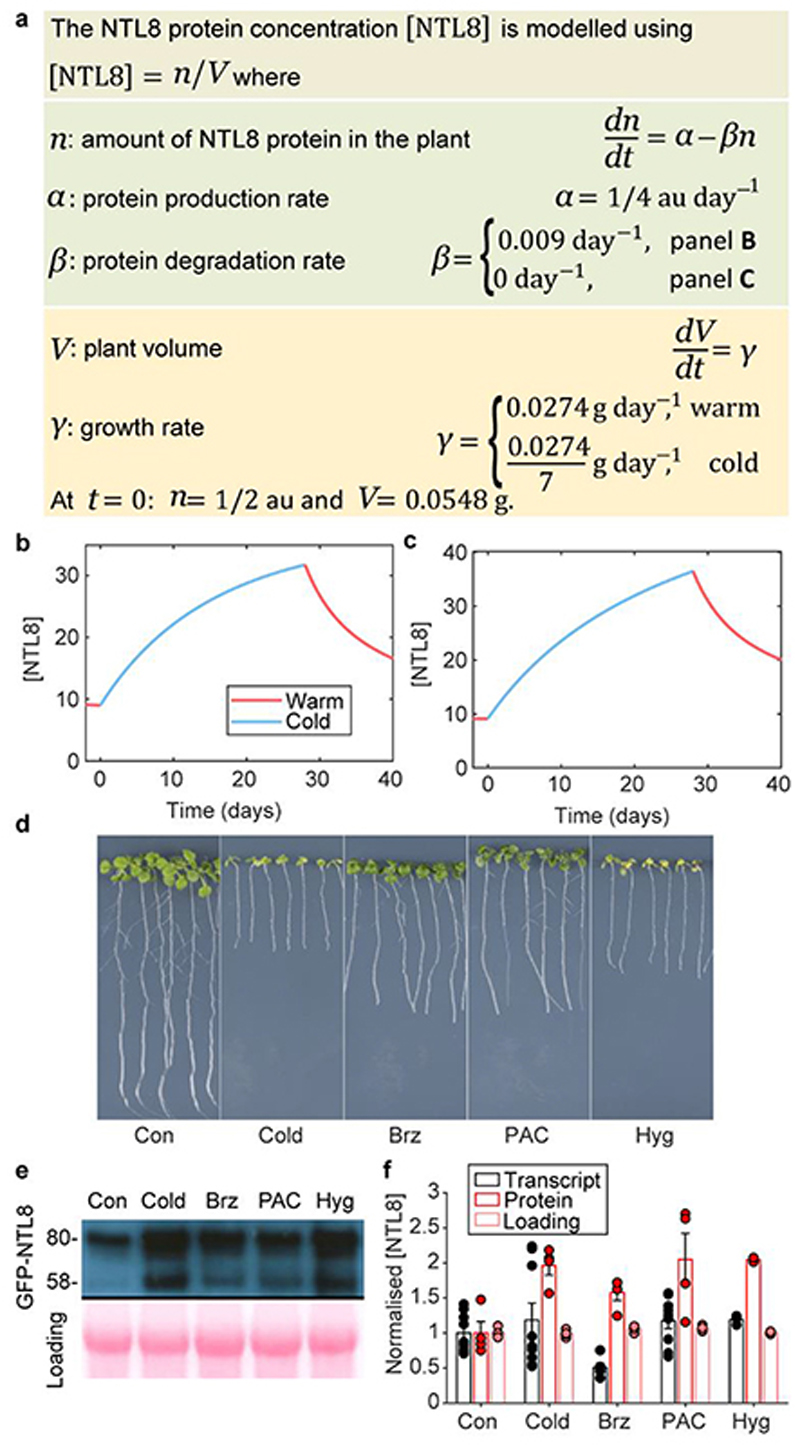
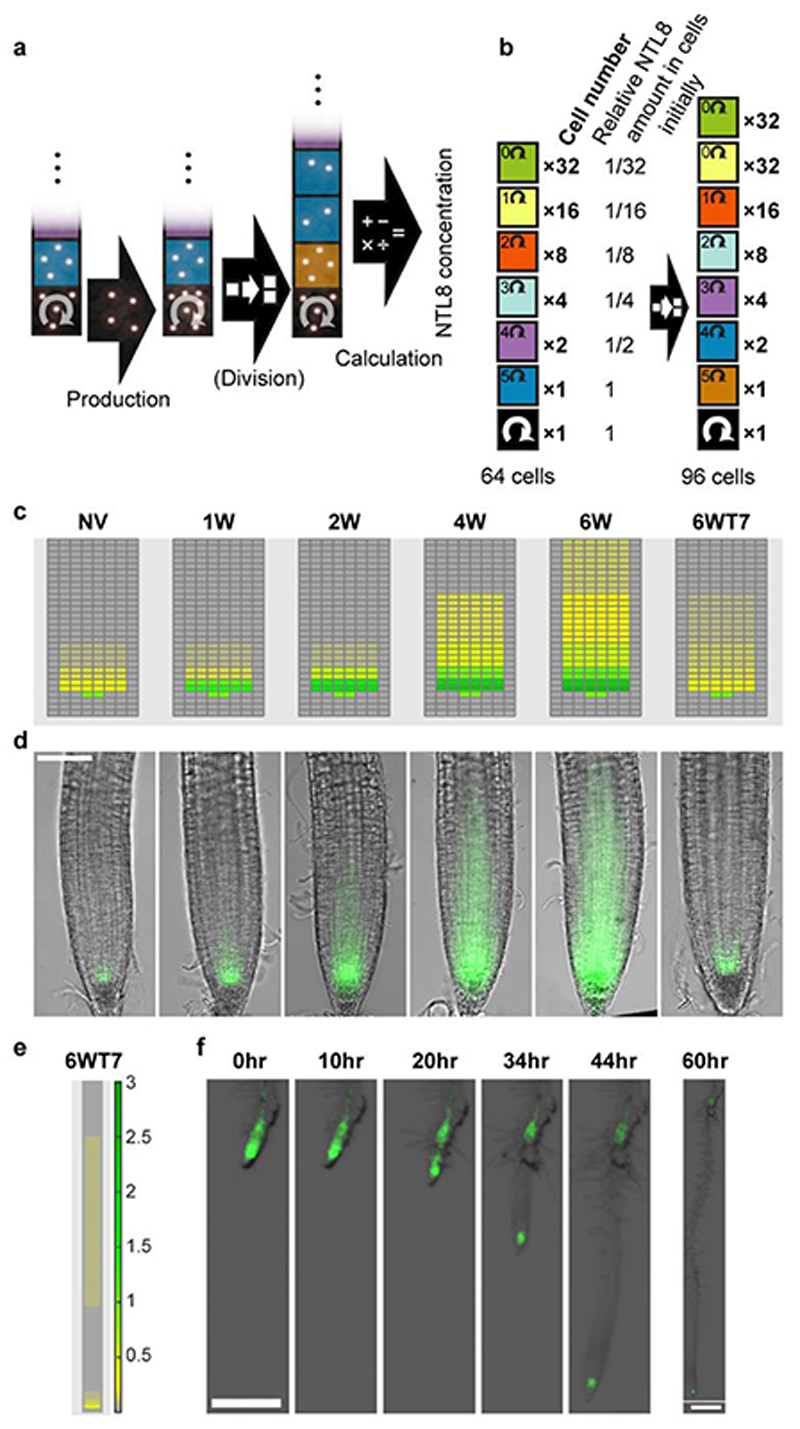
Temperature is a key factor in the growth and development of all organisms1,2. Plants have to interpret temperature fluctuations, over hourly to monthly timescales, to align their growth and development with the seasons. Much is known about how plants respond to acute thermal stresses3,4, but the mechanisms that integrate long-term temperature exposure remain unknown. The slow, winter-long upregulation of VERNALIZATION INSENSITIVE 3 (VIN3)5-7, a PHD protein that functions with Polycomb repressive complex 2 to epigenetically silence FLOWERING LOCUS C (FLC) during vernalization, is central to plants interpreting winter progression5,6,8-11. Here, by a forward genetic screen, we identify two dominant mutations of the transcription factor NTL8 that constitutively activate VIN3 expression and alter the slow VIN3 cold induction profile. In the wild type, the NTL8 protein accumulates slowly in the cold, and directly upregulates VIN3 transcription. Through combining computational simulation and experimental validation, we show that a major contributor to this slow accumulation is reduced NTL8 dilution due to slow growth at low temperatures. Temperature-dependent growth is thus exploited through protein dilution to provide the long-term thermosensory information for VIN3 upregulation. Indirect mechanisms involving temperature-dependent growth, in addition to direct thermosensing, may be widely relevant in long-term biological sensing of naturally fluctuating temperatures.
Conflict of interest statement
Figures
Comment in
-
Growth-mediated sensing of long-term cold in plants.Nature. 2020 Jul;583(7818):690-691. doi: 10.1038/d41586-020-02060-7. Nature. 2020. PMID: 32669722 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
