

Insights into epigenetic patterns in mammalian early embryos
- PMID: 32671792
- PMCID: PMC7815849
- DOI: 10.1007/s13238-020-00757-z
Insights into epigenetic patterns in mammalian early embryos
Abstract
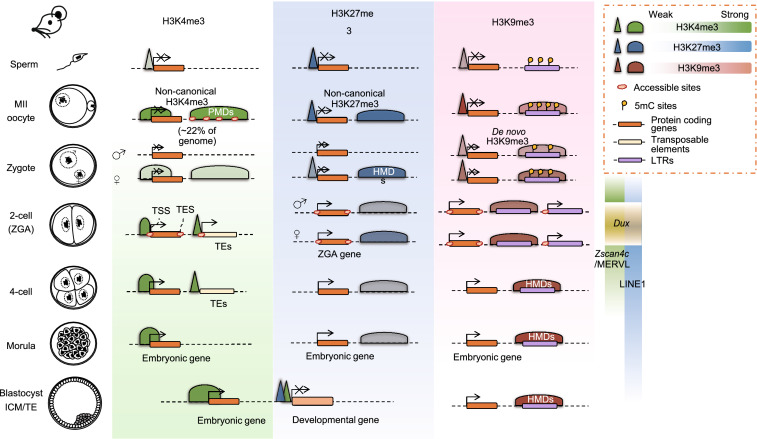
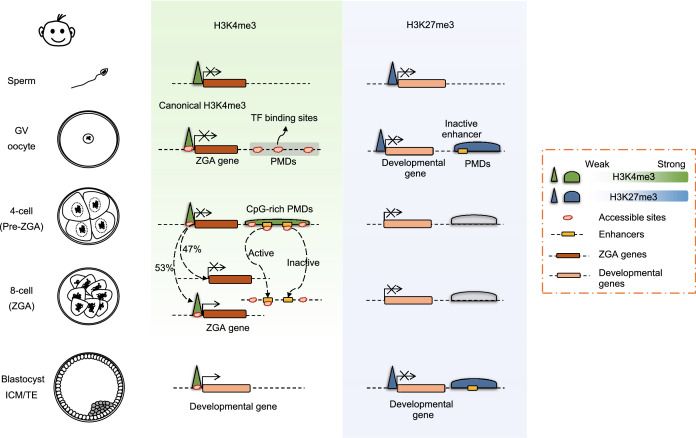
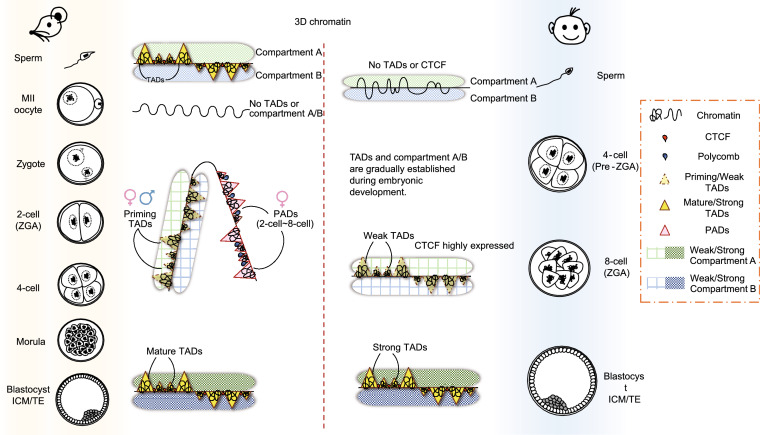
Mammalian fertilization begins with the fusion of two specialized gametes, followed by major epigenetic remodeling leading to the formation of a totipotent embryo. During the development of the pre-implantation embryo, precise reprogramming progress is a prerequisite for avoiding developmental defects or embryonic lethality, but the underlying molecular mechanisms remain elusive. For the past few years, unprecedented breakthroughs have been made in mapping the regulatory network of dynamic epigenomes during mammalian early embryo development, taking advantage of multiple advances and innovations in low-input genome-wide chromatin analysis technologies. The aim of this review is to highlight the most recent progress in understanding the mechanisms of epigenetic remodeling during early embryogenesis in mammals, including DNA methylation, histone modifications, chromatin accessibility and 3D chromatin organization.
Keywords: DNA methylation; early embryo development; epigenetic reprogramming; histone modifications.
Conflict of interest statement
Ruimin Xu, Chong Li, Xiaoyu Liu and Shaorong Gao declare that they have no conflict of interest. This article does not contain any studies with human or animal subjects performed by the any of the authors.
Figures
References
-
- Amdani SN, Yeste M, Jones C, Coward K. Sperm factors and oocyte activation: current controversies and considerations. Biol Reprod. 2015;93:50. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
