

Neutrophil Extracellular Trap Degradation by Differently Polarized Macrophage Subsets
- PMID: 32673525
- PMCID: PMC7447175
- DOI: 10.1161/ATVBAHA.120.314883
Neutrophil Extracellular Trap Degradation by Differently Polarized Macrophage Subsets
Abstract
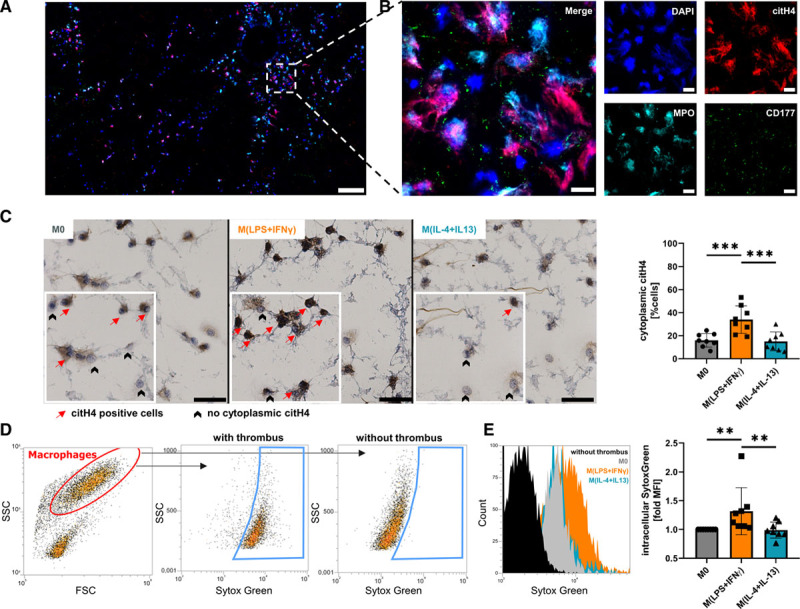
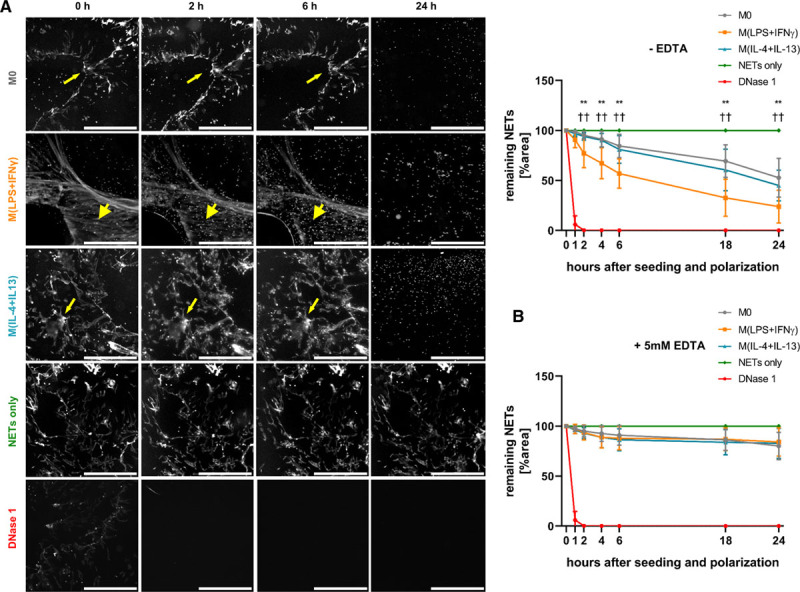
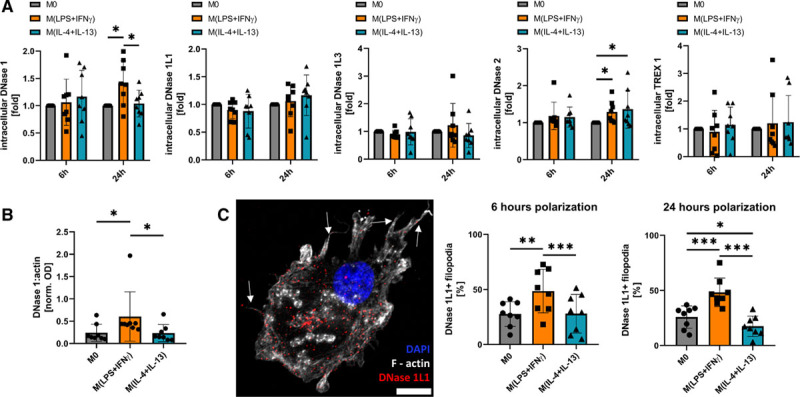
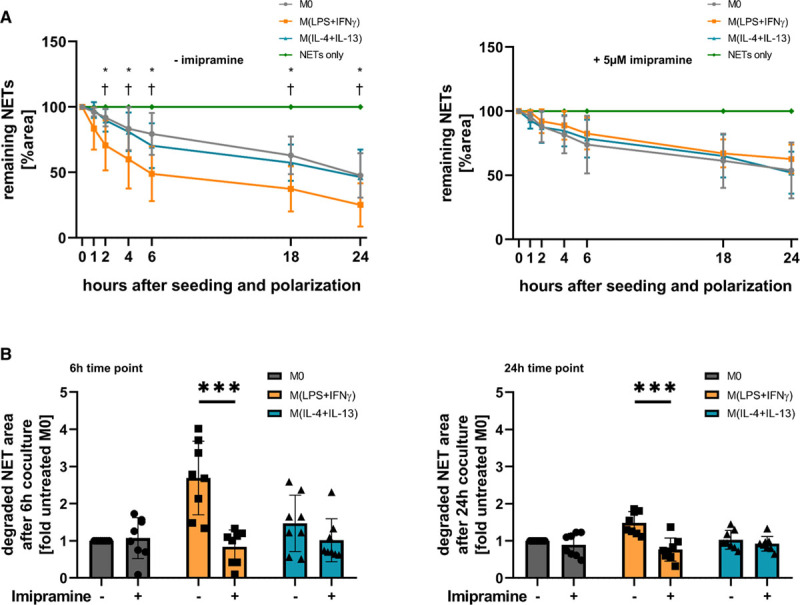
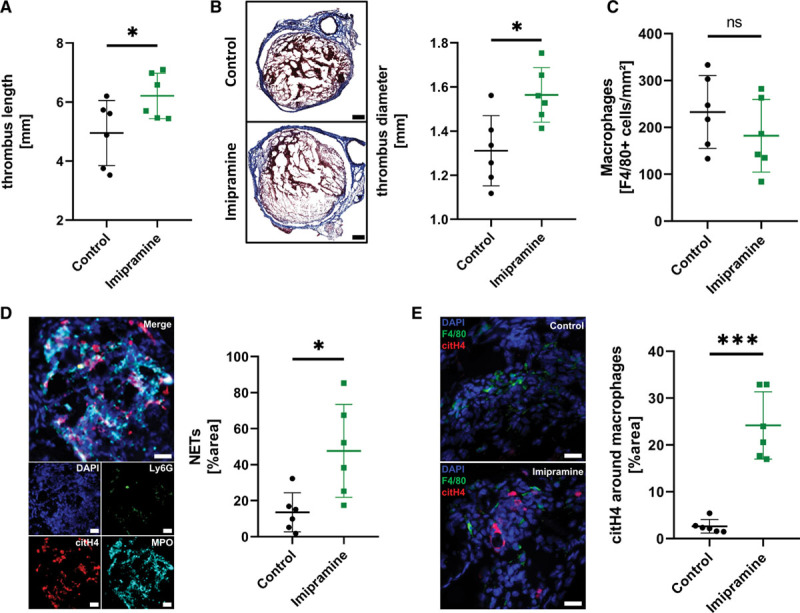
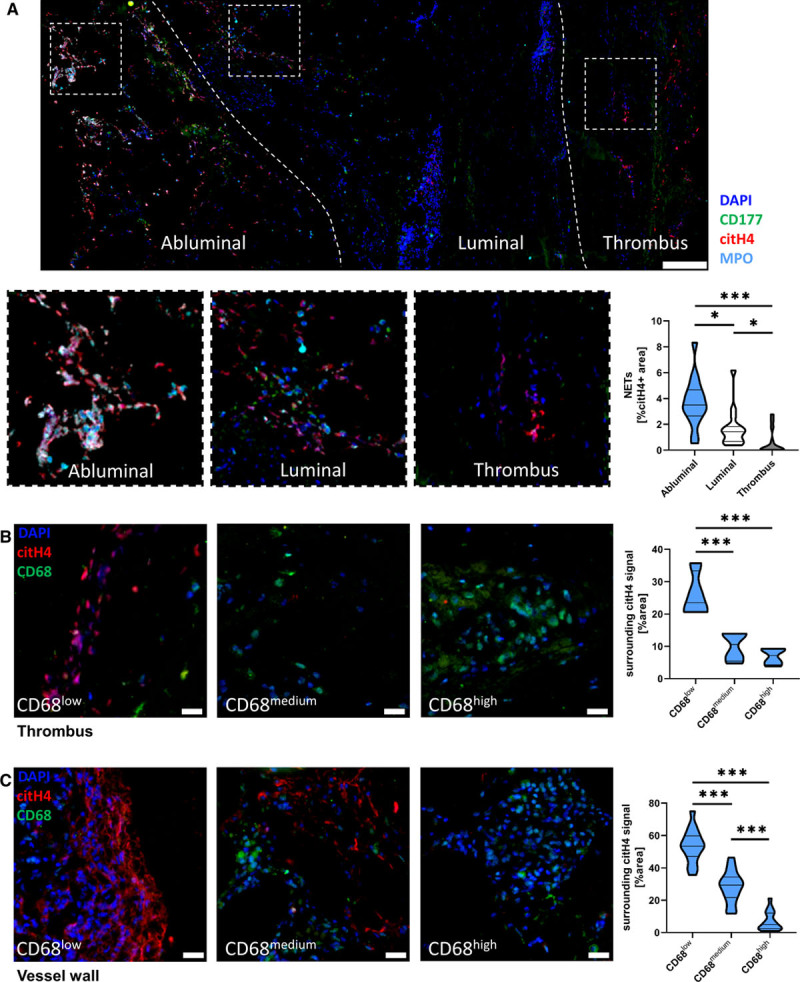
Objective: Macrophages are immune cells, capable to remodel the extracellular matrix, which can harbor extracellular DNA incorporated into neutrophil extracellular traps (NETs). To study the breakdown of NETs we studied the capability of macrophage subsets to degrade these structures in vitro and in vivo in a murine thrombosis model. Furthermore, we analyzed human abdominal aortic aneurysm samples in support of our in vitro and in vivo results. Approach and Results: Macrophages were seeded onto blood clots or isolated NETs and polarized. All macrophages were capable to degrade NETs. For initial breakdown, macrophages relied on extracellular deoxyribonucleases. Proinflammatory polarization enhanced NET degradation. The boost in degradation was because of increased macropinocytosis, as inhibition by imipramine diminished their NET breakdown. Inhibition of macropinocytosis in a murine thrombosis model led to increased NET burden and reduced thrombus resolution in vivo. When analyzing abdominal aortic aneurysm samples, macrophage density furthermore corresponded negatively with the amount of local NETs in the intraluminal thrombi as well as in the vessel wall, as increased macrophage density was associated with a reduction in NET burden.
Conclusions: We provide evidence that macrophages degrade NETs by extracellular predigestion and subsequent uptake. Furthermore, we show that proinflammatory macrophages increase NET degradation through enhanced macropinocytosis, priming them for NET engulfment. Based on our findings, that inhibition of macropinocytosis in mice corresponded to increased NET amounts in thrombi and that local macrophage density in human abdominal aortic aneurysm is negatively associated with surrounding NETs, we hypothesize, that macrophages are able to degrade NETs in vivo.
Keywords: aortic aneurysm; extracellular traps; humans; macrophages; thrombosis.
Conflict of interest statement
None.
Figures
Comment in
-
Resolving Thrombosis: Resting and Polarized Macrophages Differentially Degrade Neutrophil Extracellular Traps.Arterioscler Thromb Vasc Biol. 2020 Sep;40(9):1961-1963. doi: 10.1161/ATVBAHA.120.315013. Epub 2020 Aug 26. Arterioscler Thromb Vasc Biol. 2020. PMID: 32845774 No abstract available.
References
-
- Murray PJ. Macrophage polarization. Annu Rev Physiol 201779541–566doi: 10.1146/annurev-physiol-022516-034339 - PubMed
-
- Mantovani A, Biswas SK, Galdiero MR, Sica A, Locati M. Macrophage plasticity and polarization in tissue repair and remodelling. J Pathol 2013229176–185doi: 10.1002/path.4133 - PubMed
-
- Lim W, Gee K, Mishra S, Kumar A. Regulation of B7.1 costimulatory molecule is mediated by the IFN regulatory factor-7 through the activation of JNK in lipopolysaccharide-stimulated human monocytic cells. J Immunol 20051755690–5700doi: 10.4049/jimmunol.175.9.5690 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
