

Longitudinal Isolation of Potent Near-Germline SARS-CoV-2-Neutralizing Antibodies from COVID-19 Patients
- PMID: 32673567
- PMCID: PMC7355337
- DOI: 10.1016/j.cell.2020.06.044
Longitudinal Isolation of Potent Near-Germline SARS-CoV-2-Neutralizing Antibodies from COVID-19 Patients
Erratum in
-
Longitudinal Isolation of Potent Near-Germline SARS-CoV-2-Neutralizing Antibodies from COVID-19 Patients.Cell. 2020 Sep 17;182(6):1663-1673. doi: 10.1016/j.cell.2020.08.046. Cell. 2020. PMID: 32946786 Free PMC article. No abstract available.
Abstract
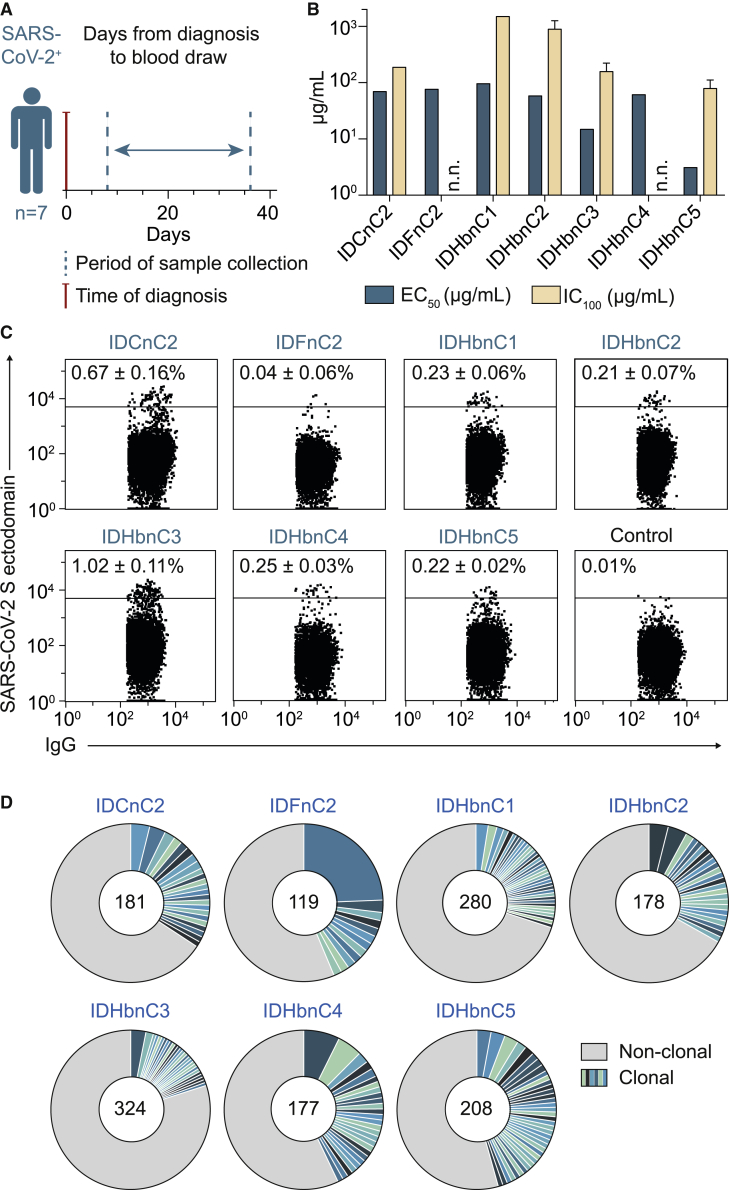
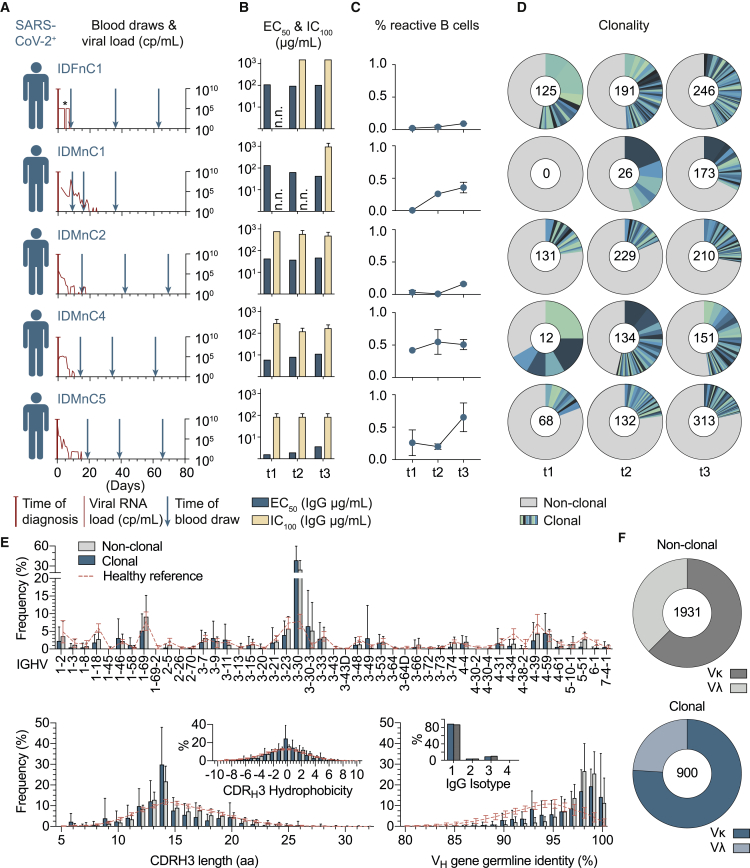
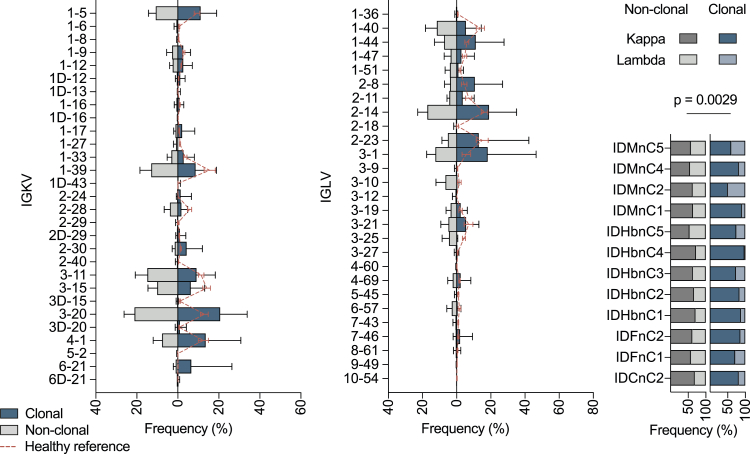
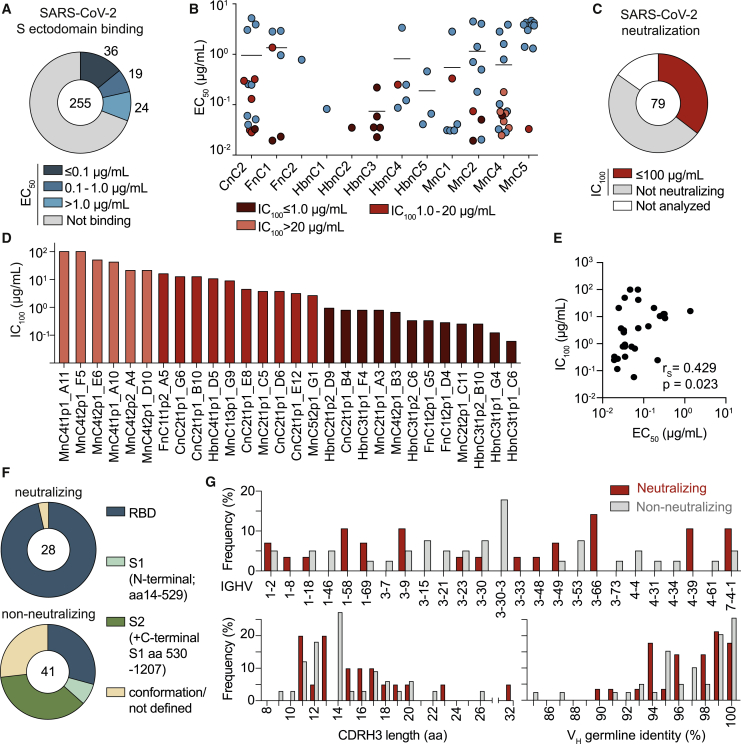
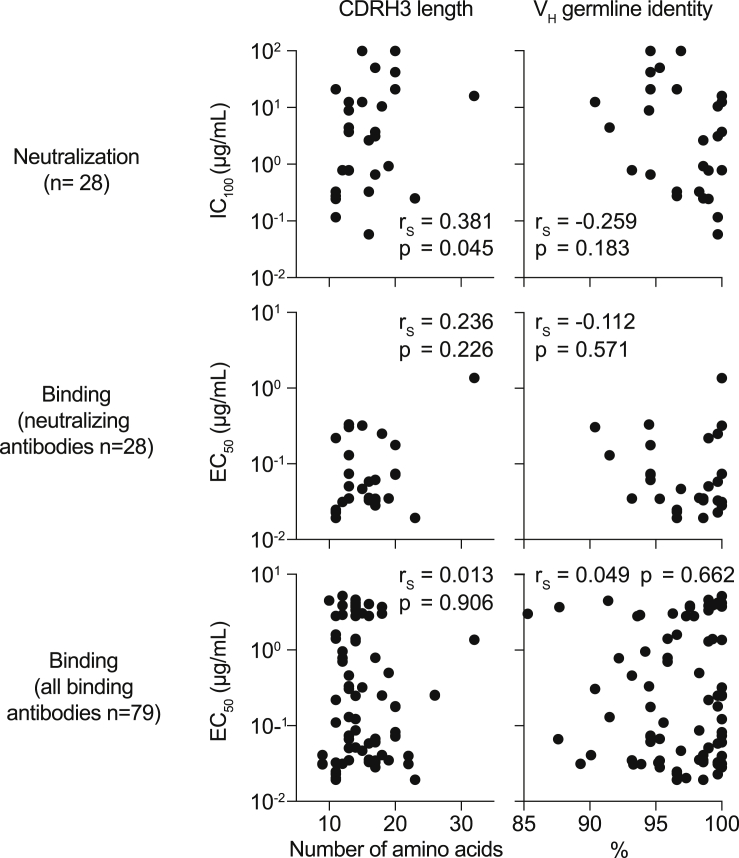
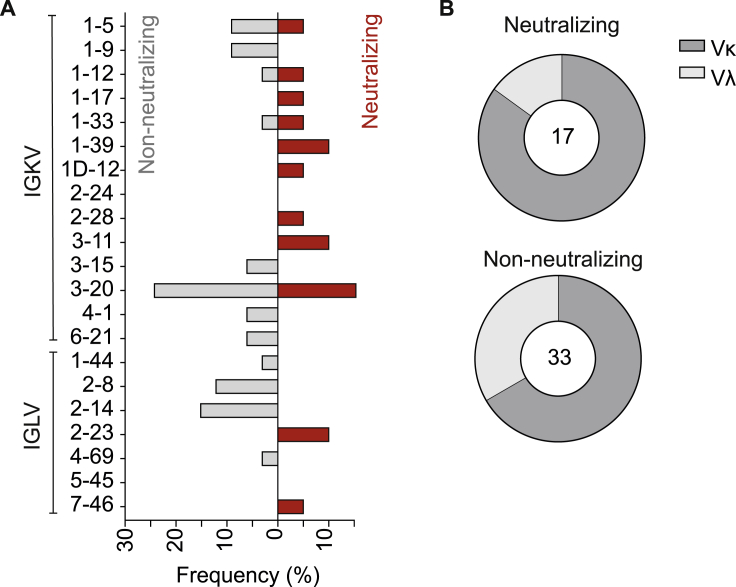
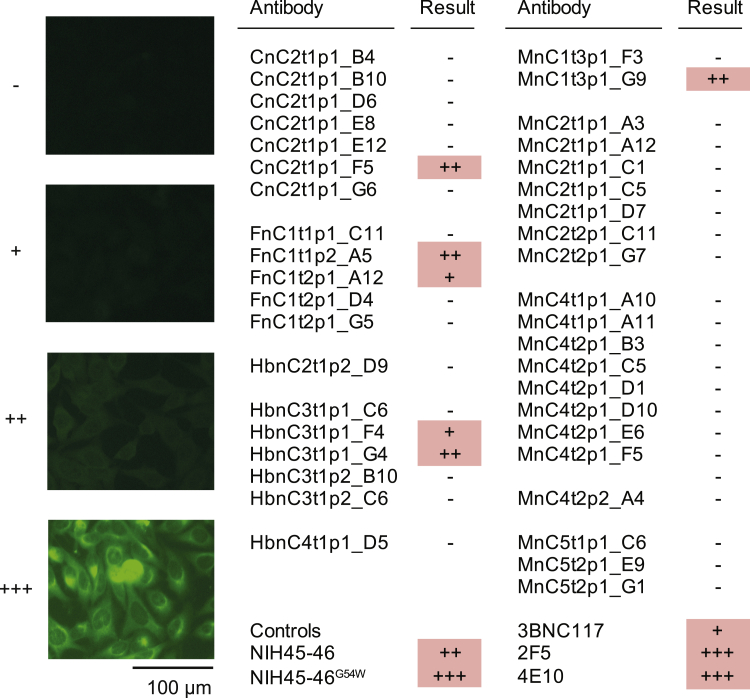
The SARS-CoV-2 pandemic has unprecedented implications for public health, social life, and the world economy. Because approved drugs and vaccines are limited or not available, new options for COVID-19 treatment and prevention are in high demand. To identify SARS-CoV-2-neutralizing antibodies, we analyzed the antibody response of 12 COVID-19 patients from 8 to 69 days after diagnosis. By screening 4,313 SARS-CoV-2-reactive B cells, we isolated 255 antibodies from different time points as early as 8 days after diagnosis. Of these, 28 potently neutralized authentic SARS-CoV-2 with IC100 as low as 0.04 μg/mL, showing a broad spectrum of variable (V) genes and low levels of somatic mutations. Interestingly, potential precursor sequences were identified in naive B cell repertoires from 48 healthy individuals who were sampled before the COVID-19 pandemic. Our results demonstrate that SARS-CoV-2-neutralizing antibodies are readily generated from a diverse pool of precursors, fostering hope for rapid induction of a protective immune response upon vaccination.
Keywords: 2019-nCoV; COVID-19; SARS-CoV-2; monoclonal antibody; neutralizing antibody; single B cell analysis.
Copyright © 2020 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests A patent application encompassing aspects of this work has been filed by the University of Cologne, listing F.K., S.B., C.K., M.Z., and H.G. as inventors.
Figures


References
-
- Brouwer P.J.M., Caniels T.G., van der Straten K., Snitselaar J.L., Aldon Y., Bangaru S., Torres J.L., Okba N.M.A., Claireaux M., Kerster G. Potent neutralizing antibodies from COVID-19 patients define multiple targets of vulnerability. Science. 2020 doi: 10.1126/science.abc5902. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
