

Review
doi: 10.1016/j.sbi.2020.06.009.
Epub 2020 Jul 13.
Structural insights into DNA loop extrusion by SMC protein complexes
Affiliations
- PMID: 32674008
- PMCID: PMC7612452
- DOI: 10.1016/j.sbi.2020.06.009
Item in Clipboard
Review
Structural insights into DNA loop extrusion by SMC protein complexes
Curr Opin Struct Biol.
2020 Dec.
Abstract
Structural Maintenance of Chromosomes (SMC) protein complexes play key roles in the three-dimensional organization of genomes in all kingdoms of life. Recent insights from chromosome contact mapping experiments and single-molecule imaging assays suggest that these complexes achieve distinct cellular functions by extruding large loops of DNA while they move along the chromatin fiber. In this short review, we summarize recent insights into the molecular architecture of these unconventional DNA motor complexes, their interaction with their DNA substrates, and the remarkable dynamic changes they can undergo during their ATPase reaction cycle.
Copyright © 2020 Elsevier Ltd. All rights reserved.
Figures
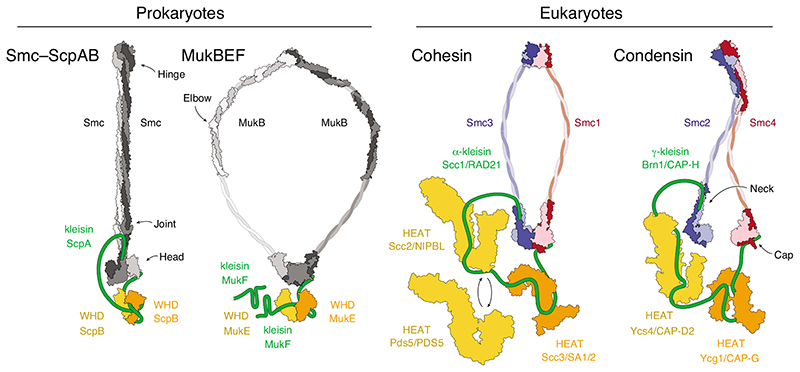
Cartoon representations of available subunit structures of prokaryotic Smc–ScpAB and MukBEF complexes and eukaryotic cohesin and condensin complexes. Available structural models are indicated for the Pyrocuccus furiosus Smc dimer (light/dark grey) in the rod-shaped, nucleotide-free state [42] and Geobacillus stearothermophilus ScpB (orange/yellow; pdb: 3W6J) [28]; the Escherichia coli MukB dimer (light/dark grey) hinge (pdb: 3IBP) [53] and coiled-coil elbow (pdb: 6H2X) [44] and Haemophilus ducreyi head domains (pdb: 3EUK) in the nucleotide-bound state and E. coli MukE (orange/yellow; pdb: 3EUH) [54]; the cohesin Smc1–Smc3 dimer (red/blue) Mus musculus hinge (pdb: 2WD5) [32] and Saccharomyces cerevisiae/Chaetomium thermophilum heads (pdb: 6QPW) in the nucleotide-bound state [31] and C. thermophilum Scc2 (yellow; pdb: 5T8V) [55], Lachancea thermotolerans Pds5 (yellow; pdb: 5F0O) [56] and Zygosaccharomyces rouxii Scc3 (orange; pdb: 4UVK) [57]; the condensin Smc2–Smc4 dimer (blue/red) S. cerevisiae hinge (pdb: 4RSI) [43] and C. thermophilum heads (pdb: 6QJ1 and 6QJ2) in the nucleotide-free state and Ycs4 (yellow; pdb: 6QJ3) [49] and S. cerevisiae Ycg1 (orange; pdb: 5OQQ) [20]. The cartoons indicate different coiled-coil conformations that are presumably accessible by all complexes. Note that MukBEF complexes can form dimers of dimers via the amino terminus of the MukF kleisin subunit, indicated by the presence of a second MukF subunit.

(A) Cartoon representations of the budding yeast cohesin Ycg1–Brn1 subcomplex bound to 18 base pairs (bp) double-stranded DNA (dsDNA) (pdb: 5OQP) [20]. A dotted line indicates the ‘safety belt’ peptide loop. (B) Cartoon model of the human cohesin SA2–SCC1 subcomplex (pdb: 6QNY) [22] bound to 19 bp dsDNA (positioned based on the structure of the yeast Scc3– Scc1 subcomplex, pdb: 6H8Q) [21] and the zinc-fingers 2–7 of human CTCF bound to 23 bp dsDNA (pdb 5T0U) [58]. A dotted line indicates a linker to a peptide motif that binds to the ‘conserved essential surface’ region on the backside of SA2.
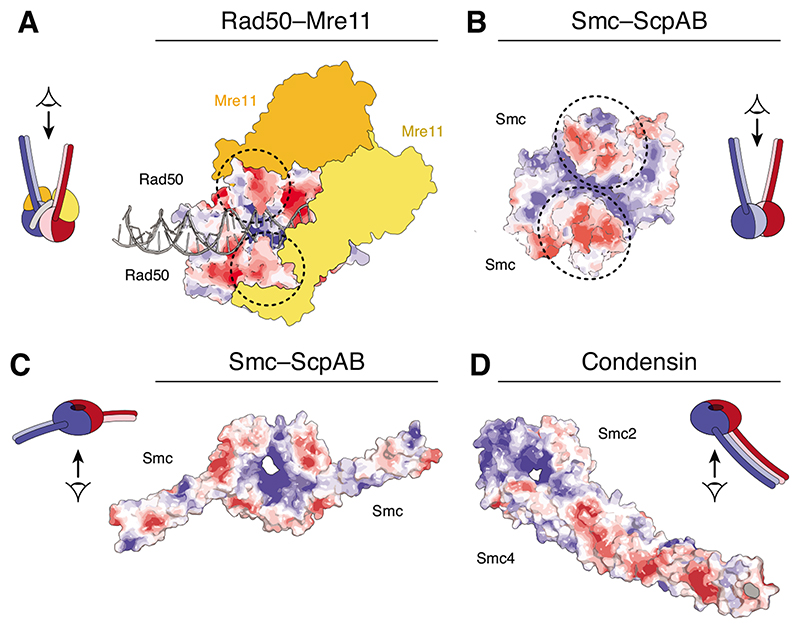
Surface charge potential representations of (A) Rad50 homodimer heads bound to 31 bp dsDNA and the Mre11 dimer (yellow/orange) (pdb: 6S85) [25], (B)
Geobacillus stearothermophilus SMC head homodimer (pdb 5H68) [59] and (C) a chimeric Saccharomyces cerevisiae/Chaetomium thermophilum Smc1–Smc3 head heterodimer (pdb: 6QPW) [31] viewed from in-between the coiled coils. Surface charge scale ranges from – 5 kT/e (red) to +5 kT/e (blue). Circles indicate the positions of the coiled-coil stems.
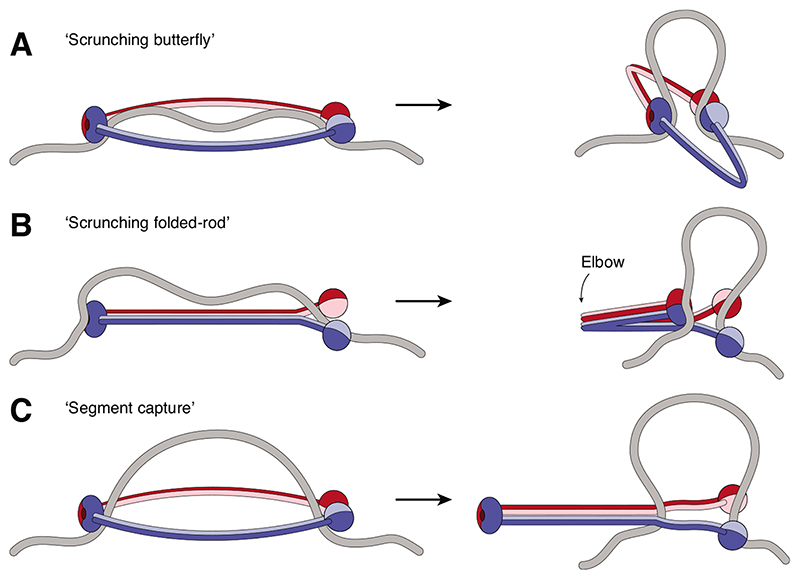
(A) Extended ring-shaped SMCs might bind to DNA via their head and hinge domains and DNA loops might be formed when the coiled-coils bend. (B) Rod-shaped SMCs might bind DNA as in (A) and then fold at their elbow region to create and extend a DNA loop. (C) Ring-shaped SMC complexes might capture a DNA loop within their lumen and then zip up their coiled coils from hinge to heads to move the loop into a new compartment created by the kleisin subunit (not depicted). In all models, the bases of the DNA loop would need to be held in place when the reaction cycle resets.
References
-
- Yatskevich S, Rhodes J, Nasmyth K. Organization of Chromosomal DNA by SMC Complexes. Annu Rev Genet. 2019;53:445–482. - PubMed
-
- Nishiyama T. Cohesion and cohesin-dependent chromatin organization. Curr Opin Cell Biol. 2019;58:8–14. - PubMed
-
- Aragon L. The Smc5/6 Complex: New and Old Functions of the Enigmatic Long-Distance Relative. Annu Rev Genet. 2018;52:89–107. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
