

Debulking of topoisomerase DNA-protein crosslinks (TOP-DPC) by the proteasome, non-proteasomal and non-proteolytic pathways
- PMID: 32674013
- PMCID: PMC9210512
- DOI: 10.1016/j.dnarep.2020.102926
Debulking of topoisomerase DNA-protein crosslinks (TOP-DPC) by the proteasome, non-proteasomal and non-proteolytic pathways
Abstract
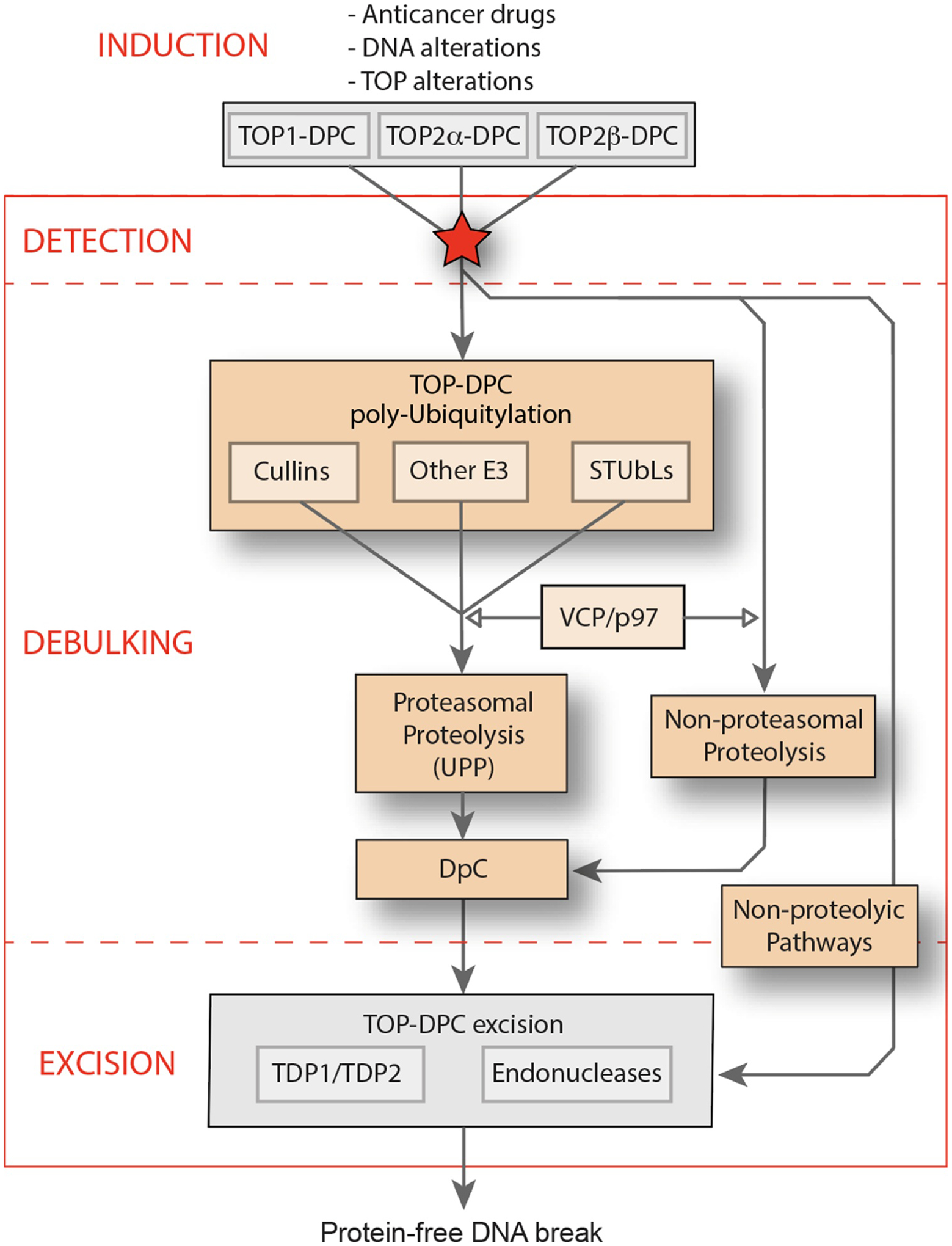
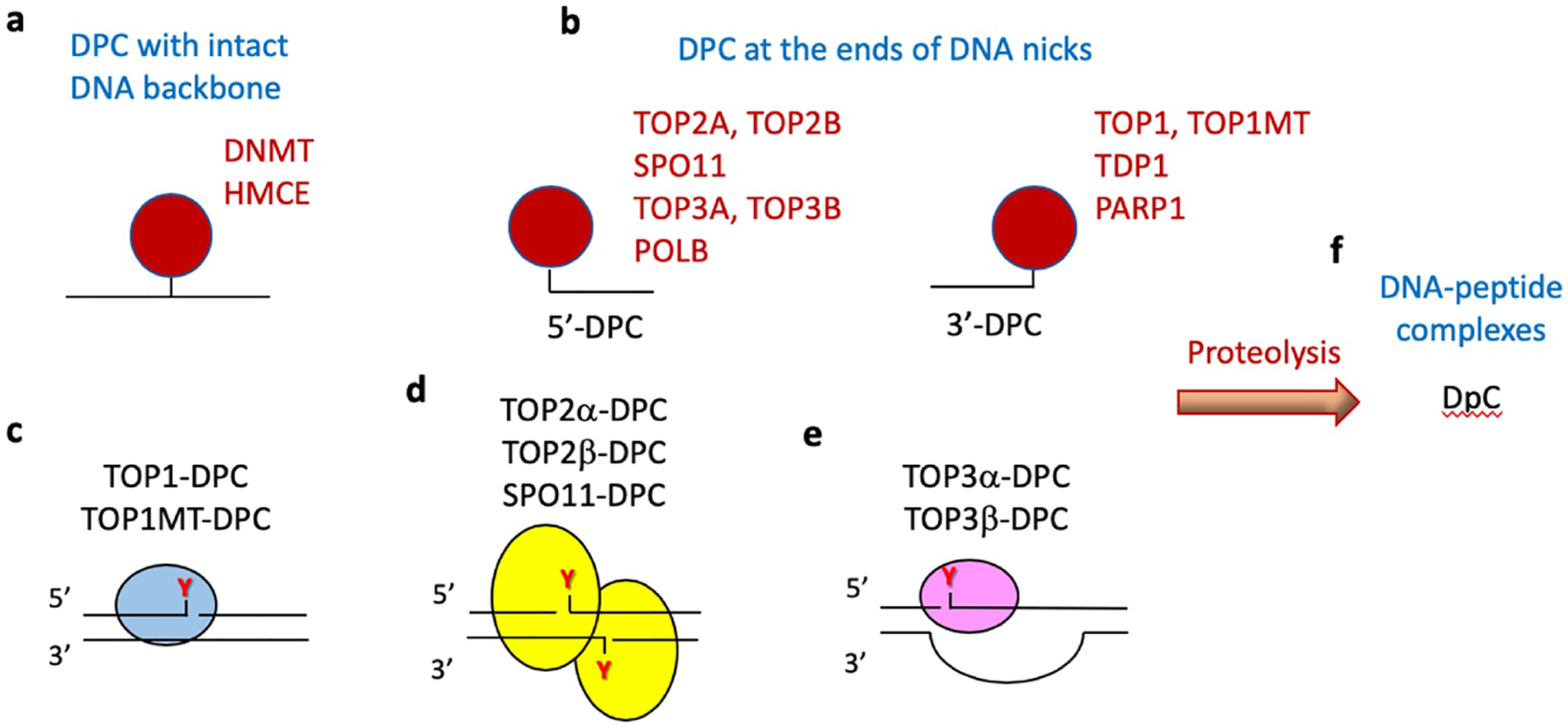
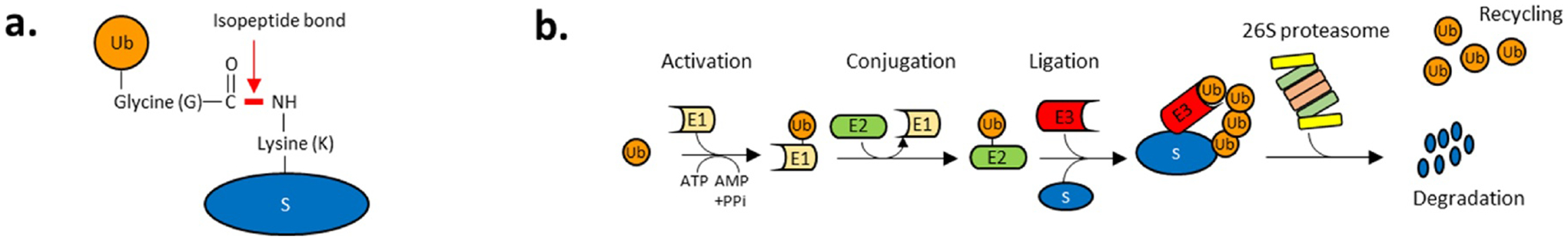
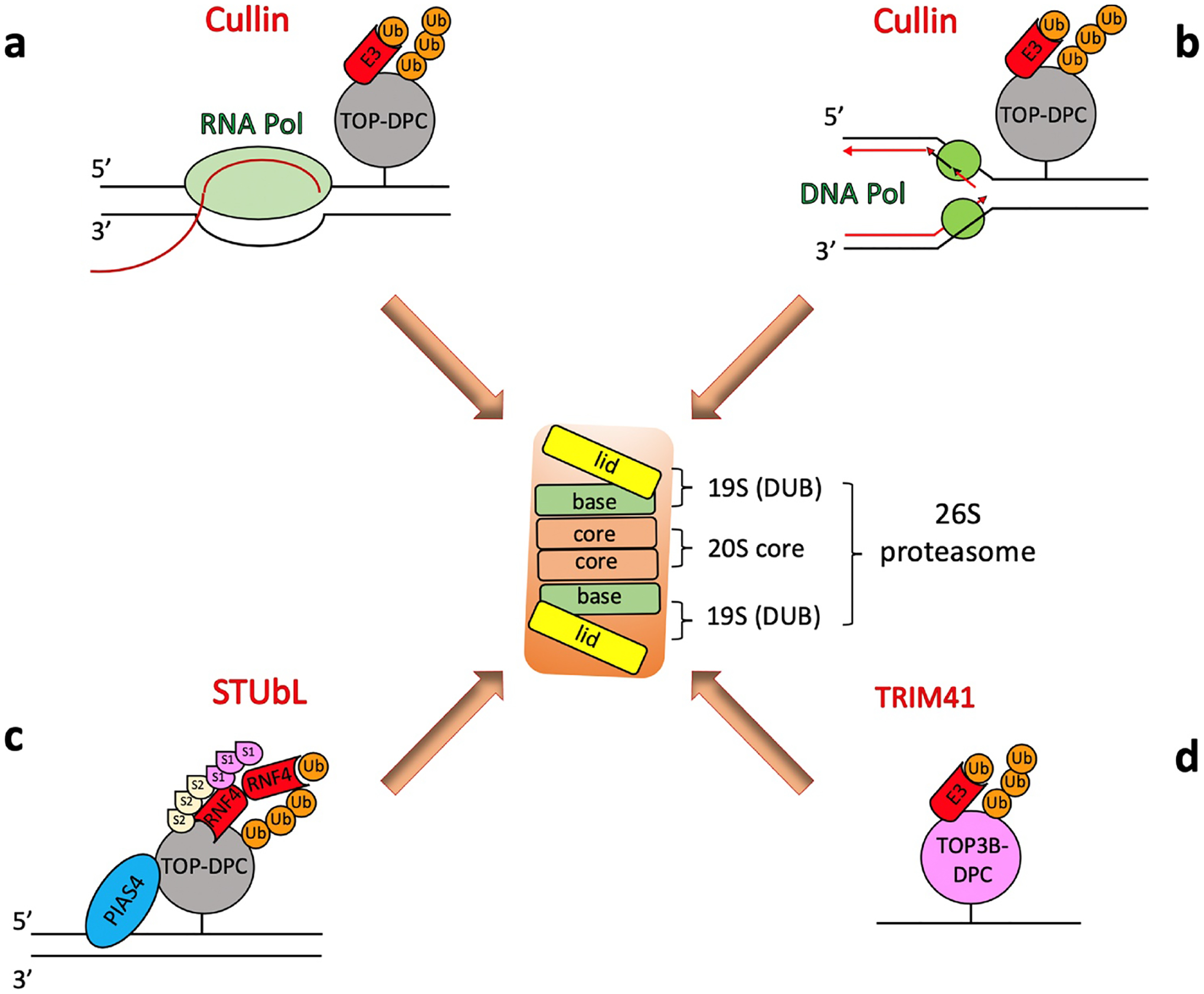
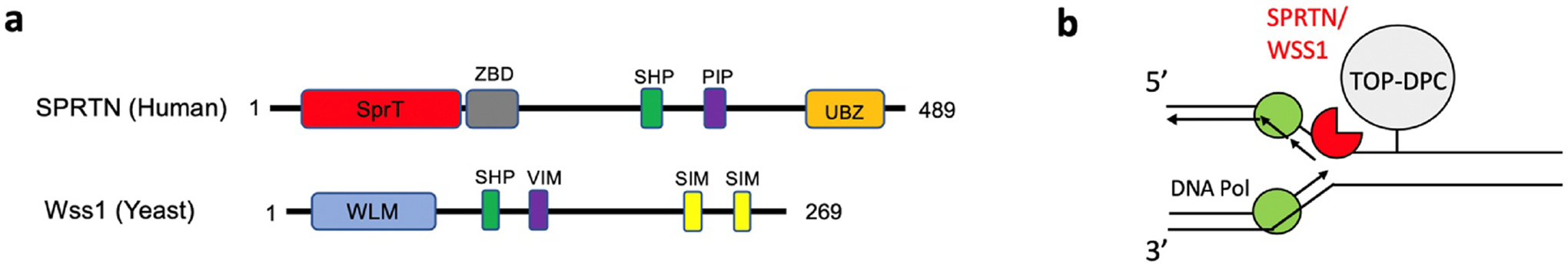
Topoisomerases play a pivotal role in ensuring DNA metabolisms during replication, transcription and chromosomal segregation. To manage DNA topology, topoisomerases generate break(s) in the DNA backbone by forming transient enzyme-DNA cleavage complexes (TOPcc) with phosphotyrosyl linkages between DNA ends and topoisomerase catalytic tyrosyl residues. Topoisomerases have been identified as the cellular targets of a variety of anti-cancer drugs (e.g. topotecan, irinotecan, etoposide and doxorubicin, and antibiotics (e.g. ciprofloxacin and levofloxacin). These drugs, as well as other exogenous and endogenous agents, convert the transient TOPcc into persistent TOPcc, which we refer to as topoisomerase DNA-protein crosslinks (TOP-DPC) that challenge genome integrity and lead to cell death if left unrepaired. Proteolysis of the bulky protein component of TOP-DPC (debulking) is a poorly understood repair process employed across eukaryotes. TOP-DPC proteolysis can be achieved either by the ubiquitin-proteasome pathway (UPP) or by non-proteasomal proteases, which are typified by the metalloprotease SPRTN/WSS1. Debulking of TOP-DPC exposes the phosphotyrosyl bonds, hence enables tyrosyl-DNA phosphodiesterases (TDP1 and TDP2) to access and cleave the bonds. In this review, we focus on current knowledge of the protease pathways for debulking TOP-DPC and highlighting recent advances in understanding the mechanisms regulating the proteolytic repair pathways. We also discuss the avenues that are being exploited to target the proteolytic repair pathways for improving the clinical outcome of topoisomerase inhibitors.
Keywords: DNA-protein crosslinks; SPRTN; SUMOylation; Topoisomerases; Ubiquitin-proteasome pathway.
Published by Elsevier B.V.
Conflict of interest statement
Declaration of Competing Interest
The authors declare that there is no conflict of interest.
Figures
Similar articles
-
Excision repair of topoisomerase DNA-protein crosslinks (TOP-DPC).DNA Repair (Amst). 2020 May;89:102837. doi: 10.1016/j.dnarep.2020.102837. Epub 2020 Mar 7. DNA Repair (Amst). 2020. PMID: 32200233 Free PMC article. Review.
-
From the TOP: Formation, recognition and resolution of topoisomerase DNA protein crosslinks.DNA Repair (Amst). 2024 Oct;142:103751. doi: 10.1016/j.dnarep.2024.103751. Epub 2024 Aug 16. DNA Repair (Amst). 2024. PMID: 39180935 Review.
-
Tyrosyl-DNA phosphodiesterase 2 (TDP2) repairs topoisomerase 1 DNA-protein crosslinks and 3'-blocking lesions in the absence of tyrosyl-DNA phosphodiesterase 1 (TDP1).DNA Repair (Amst). 2020 Jul-Aug;91-92:102849. doi: 10.1016/j.dnarep.2020.102849. Epub 2020 May 17. DNA Repair (Amst). 2020. PMID: 32460231
-
Untangling trapped topoisomerases with tyrosyl-DNA phosphodiesterases.DNA Repair (Amst). 2020 Oct;94:102900. doi: 10.1016/j.dnarep.2020.102900. Epub 2020 Jun 18. DNA Repair (Amst). 2020. PMID: 32653827 Review.
-
Tyrosyl-DNA phosphodiesterase 1 (TDP1) and SPRTN protease repair histone 3 and topoisomerase 1 DNA-protein crosslinks in vivo.Open Biol. 2023 Oct;13(10):230113. doi: 10.1098/rsob.230113. Epub 2023 Oct 4. Open Biol. 2023. PMID: 37788708 Free PMC article.
Cited by
-
TDP1 suppresses chromosomal translocations and cell death induced by abortive TOP1 activity during gene transcription.Nat Commun. 2023 Nov 9;14(1):6940. doi: 10.1038/s41467-023-42622-7. Nat Commun. 2023. PMID: 37945566 Free PMC article.
-
Reconsidering the mechanisms of action of PARP inhibitors based on clinical outcomes.Cancer Sci. 2022 Sep;113(9):2943-2951. doi: 10.1111/cas.15477. Epub 2022 Jul 16. Cancer Sci. 2022. PMID: 35766436 Free PMC article. Review.
-
Mechanisms and Regulation of DNA-Protein Crosslink Repair During DNA Replication by SPRTN Protease.Front Mol Biosci. 2022 Jun 15;9:916697. doi: 10.3389/fmolb.2022.916697. eCollection 2022. Front Mol Biosci. 2022. PMID: 35782873 Free PMC article. Review.
-
Small molecule microarray identifies inhibitors of tyrosyl-DNA phosphodiesterase 1 that simultaneously access the catalytic pocket and two substrate binding sites.Chem Sci. 2021 Jan 28;12(11):3876-3884. doi: 10.1039/d0sc05411a. Chem Sci. 2021. PMID: 34163656 Free PMC article.
-
Towards a comprehensive view of 8-oxo-7,8-dihydro-2'-deoxyguanosine: Highlighting the intertwined roles of DNA damage and epigenetics in genomic instability.DNA Repair (Amst). 2021 Jan;97:103027. doi: 10.1016/j.dnarep.2020.103027. Epub 2020 Nov 26. DNA Repair (Amst). 2021. PMID: 33285475 Free PMC article. Review.
References
-
- Stingele J, Bellelli R, Boulton SJ, Mechanisms of DNA-protein crosslink repair, Nat. Rev. Mol. Cell Biol 18 (2017) 563–573. - PubMed
-
- Ide H, Nakano T, Salem AMH, Shoulkamy MI, DNA-protein cross-links: formidable challenges to maintaining genome integrity, DNA Repair (Amst) 71 (2018) 190–197. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
