

The novel features of Plantago ovata seed mucilage accumulation, storage and release
- PMID: 32678191
- PMCID: PMC7366641
- DOI: 10.1038/s41598-020-68685-w
The novel features of Plantago ovata seed mucilage accumulation, storage and release
Abstract
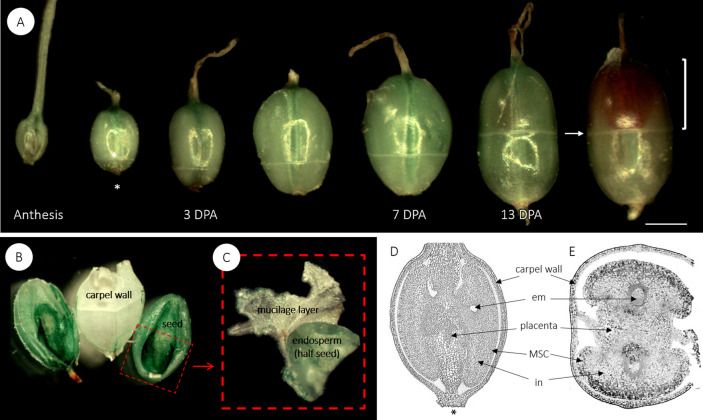
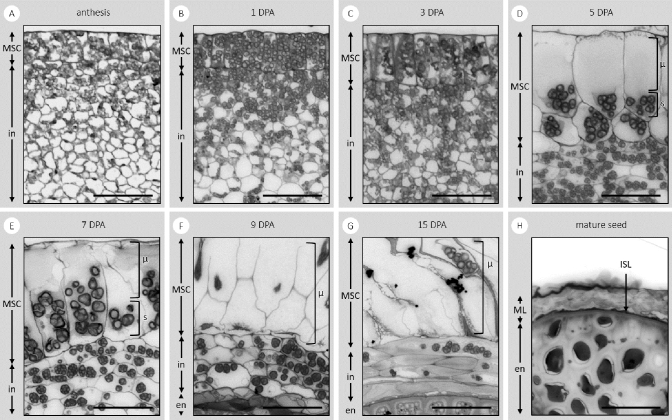
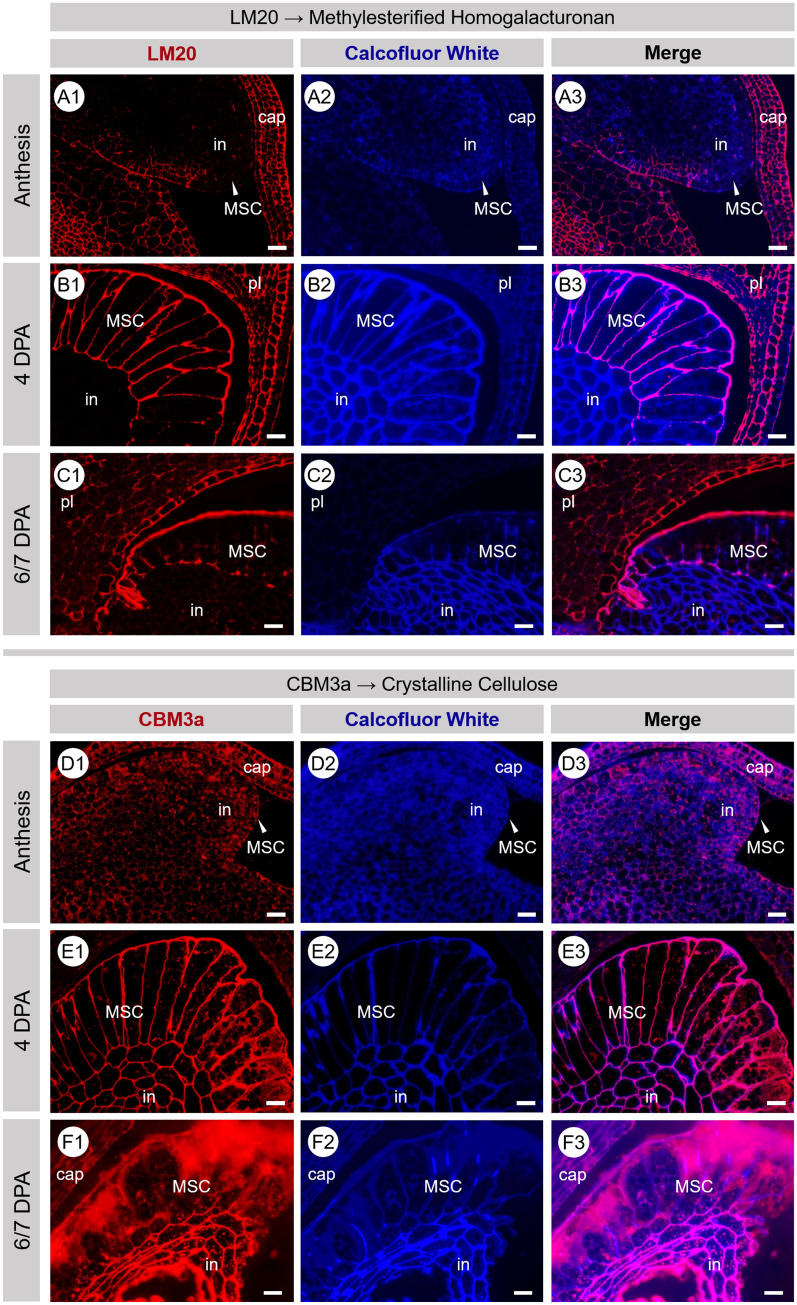
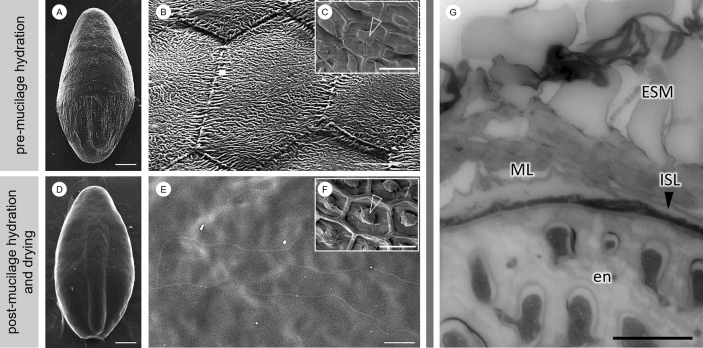
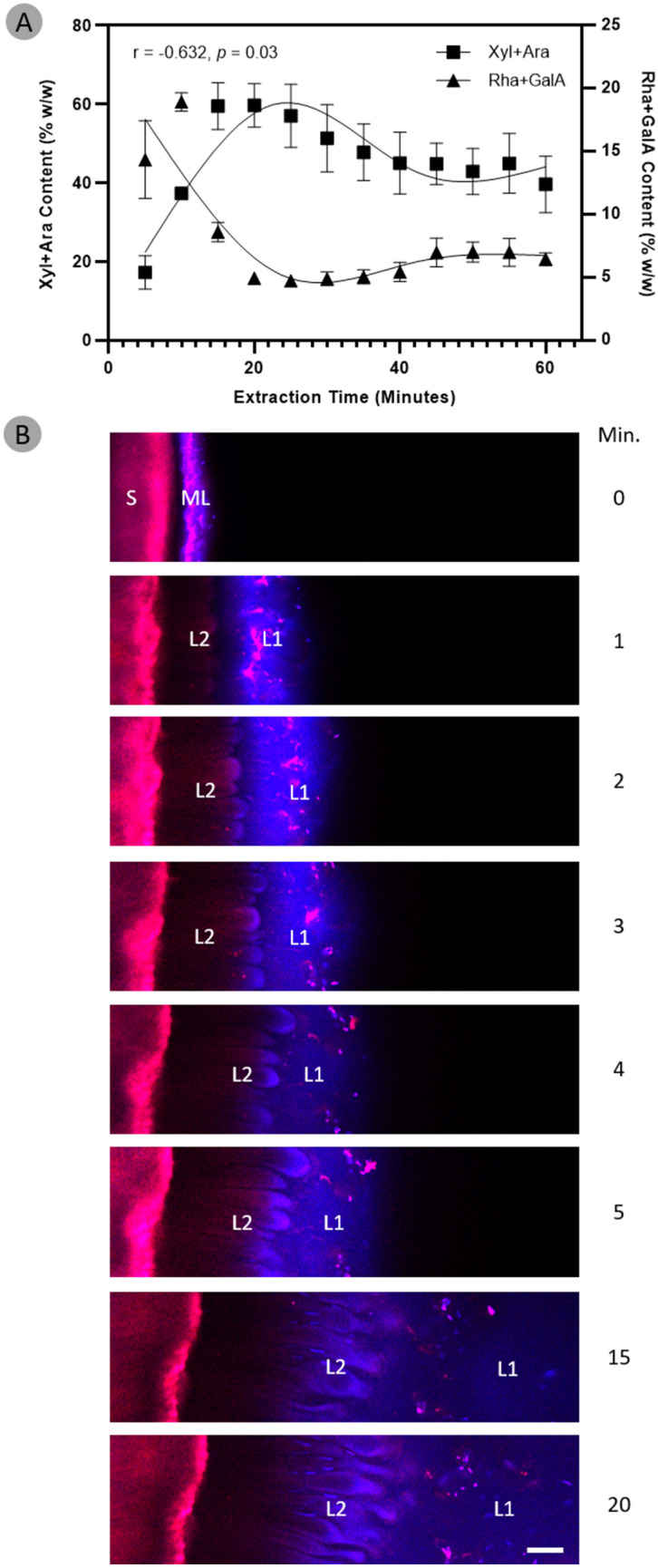
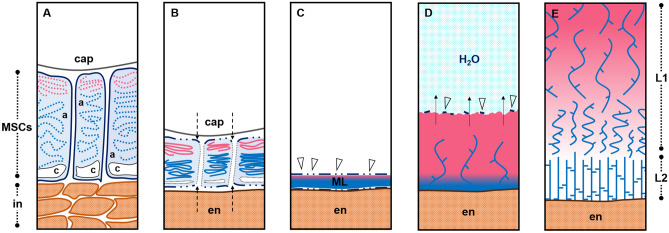
Seed mucilage polysaccharide production, storage and release in Plantago ovata is strikingly different to that of the model plant Arabidopsis. We have used microscopy techniques to track the development of mucilage secretory cells and demonstrate that mature P. ovata seeds do not have an outer intact cell layer within which the polysaccharides surround internal columellae. Instead, dehydrated mucilage is spread in a thin homogenous layer over the entire seed surface and upon wetting expands directly outwards, away from the seed. Observing mucilage expansion in real time combined with compositional analysis allowed mucilage layer definition and the roles they play in mucilage release and architecture upon hydration to be explored. The first emergent layer of hydrated mucilage is rich in pectin, extremely hydrophilic, and forms an expansion front that functions to 'jumpstart' hydration and swelling of the second layer. This next layer, comprising the bulk of the expanded seed mucilage, is predominantly composed of heteroxylan and appears to provide much of the structural integrity. Our results indicate that the synthesis, deposition, desiccation, and final storage position of mucilage polysaccharides must be carefully orchestrated, although many of these processes are not yet fully defined and vary widely between myxospermous plant species.
Figures

References
-
- Phan JL, Burton RA. New insights into the composition and structure of seed mucilage. Annu. Plant Rev. Online. 2018;1:1–41.
-
- Macquet A, Ralet M-C, Kronenberger J, Marion-Poll A, North HM. Insitu, chemical and macromolecular study of the composition of Arabidopsisthaliana seed coat mucilage. Plant Cell Physiol. 2007;48:984–999. - PubMed
-
- Harpaz-Saad S, et al. Cellulose synthesis via the FEI2 RLK/SOS5 pathway and CELLULOSE SYNTHASE 5 is required for the structure of seed coat mucilage in Arabidopsis. Plant J. 2011;68:941–953. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
