

Cell Types Promoting Goosebumps Form a Niche to Regulate Hair Follicle Stem Cells
- PMID: 32679029
- PMCID: PMC7540726
- DOI: 10.1016/j.cell.2020.06.031
Cell Types Promoting Goosebumps Form a Niche to Regulate Hair Follicle Stem Cells
Abstract
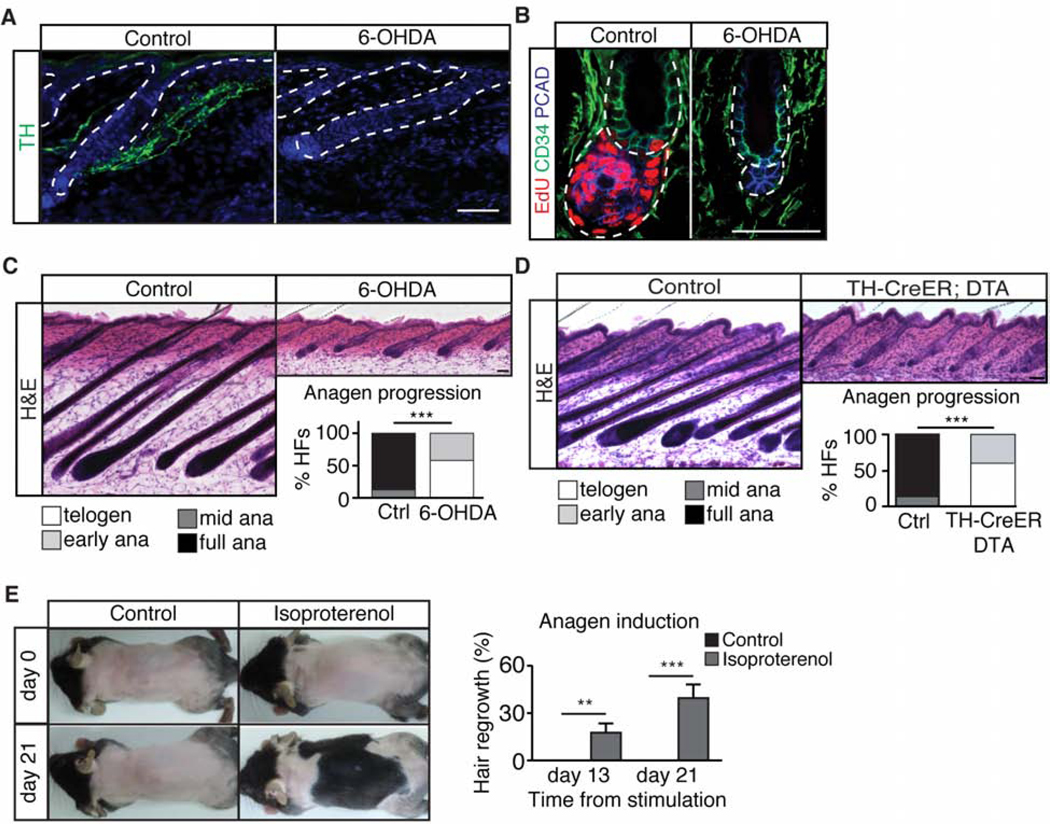
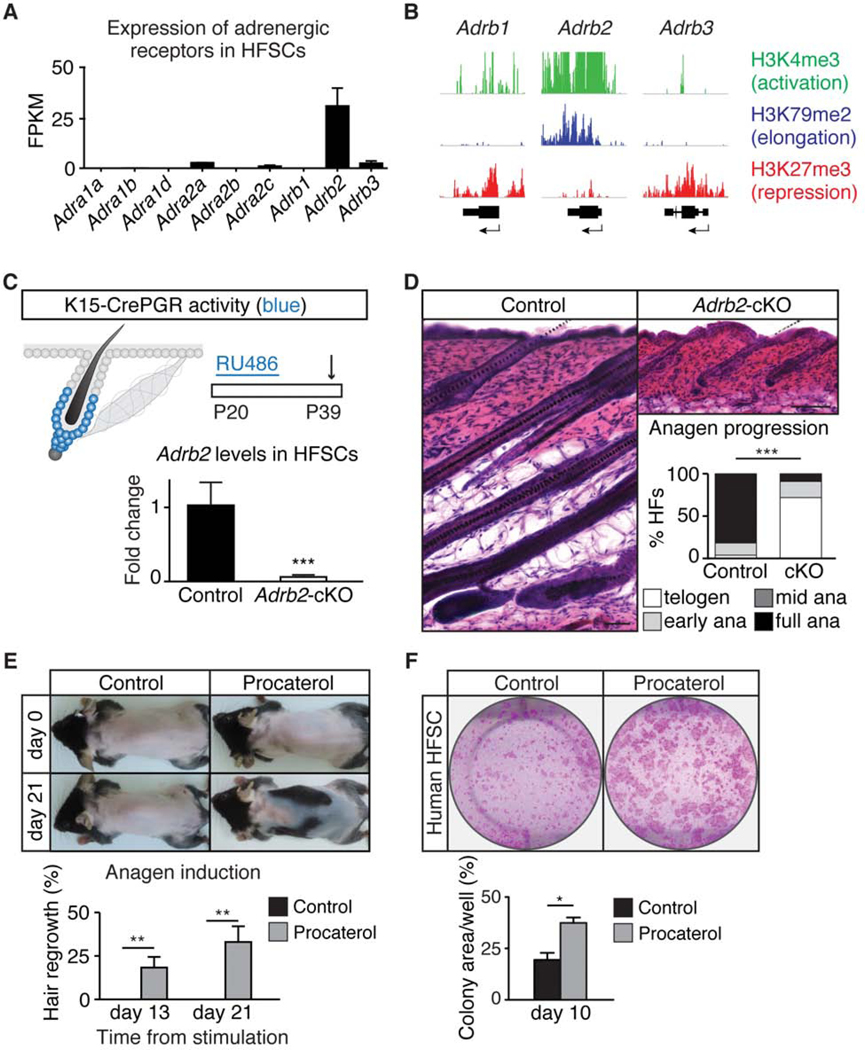
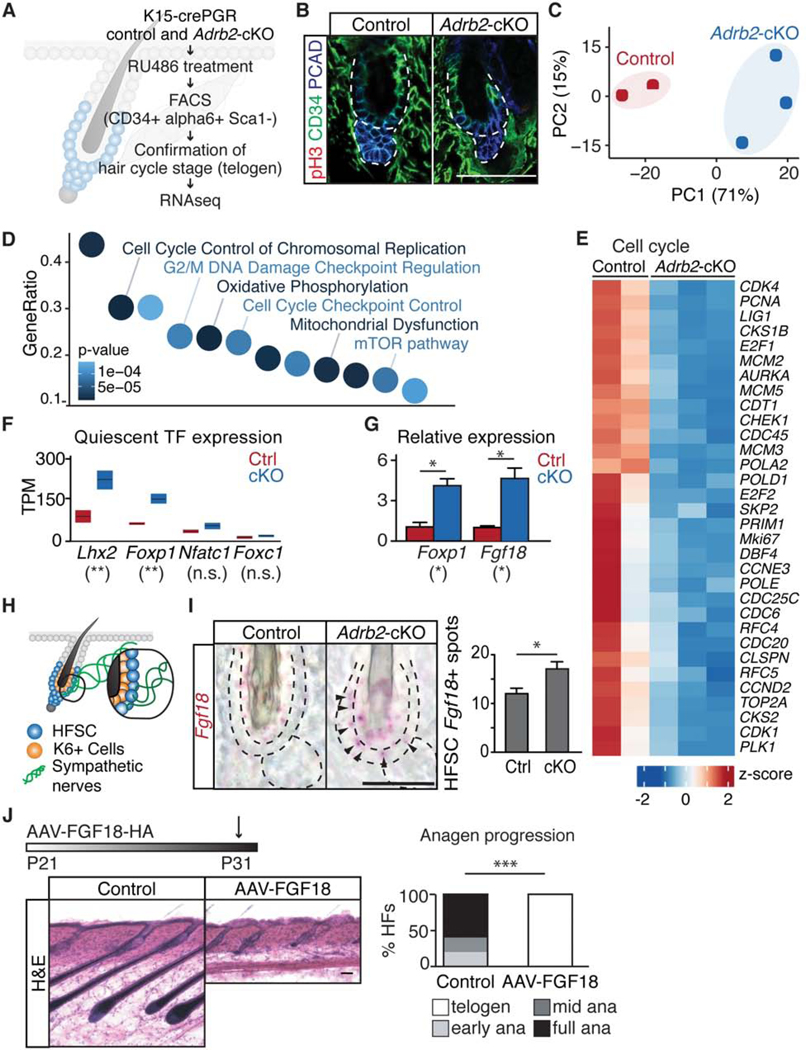
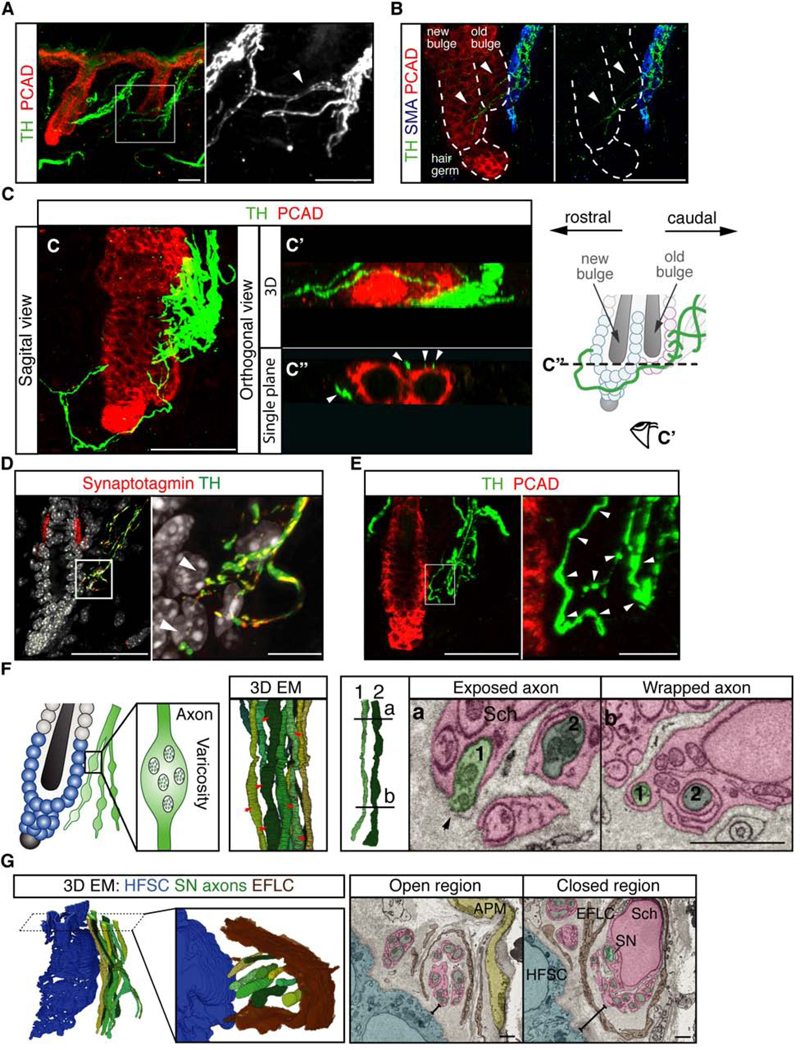
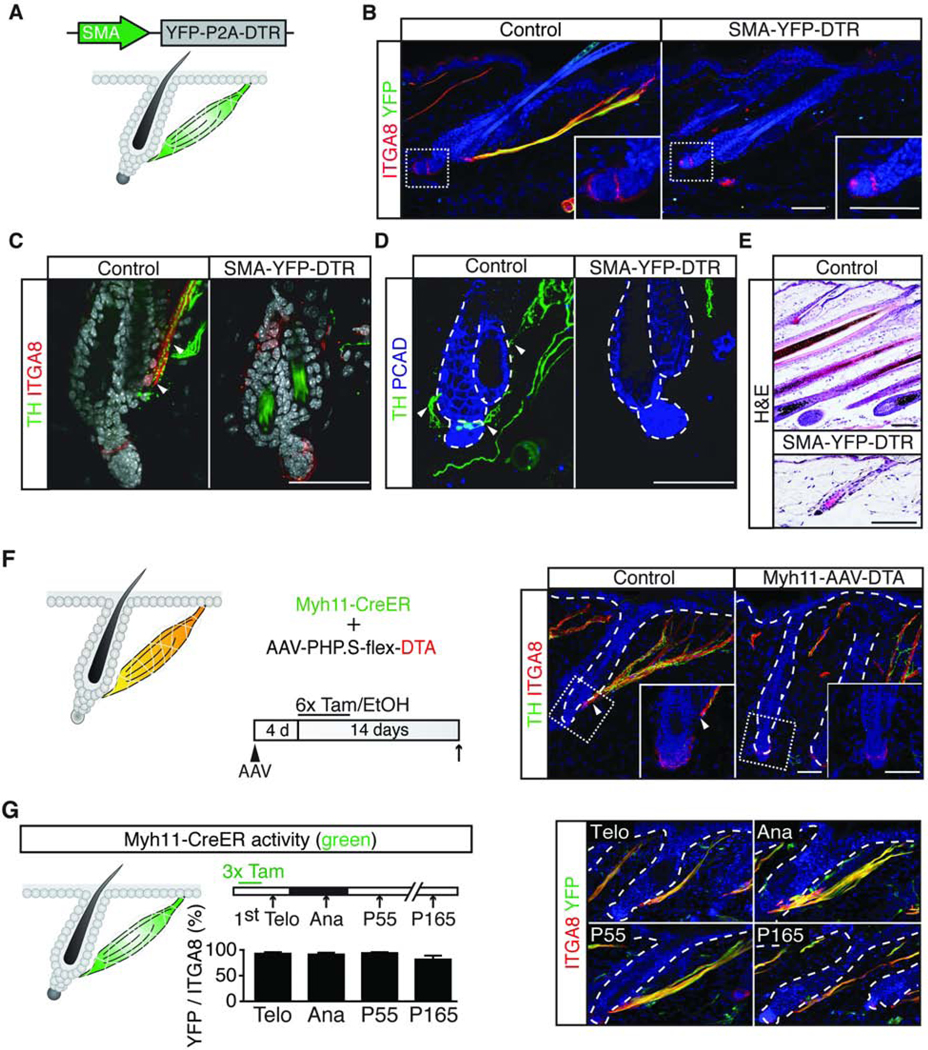
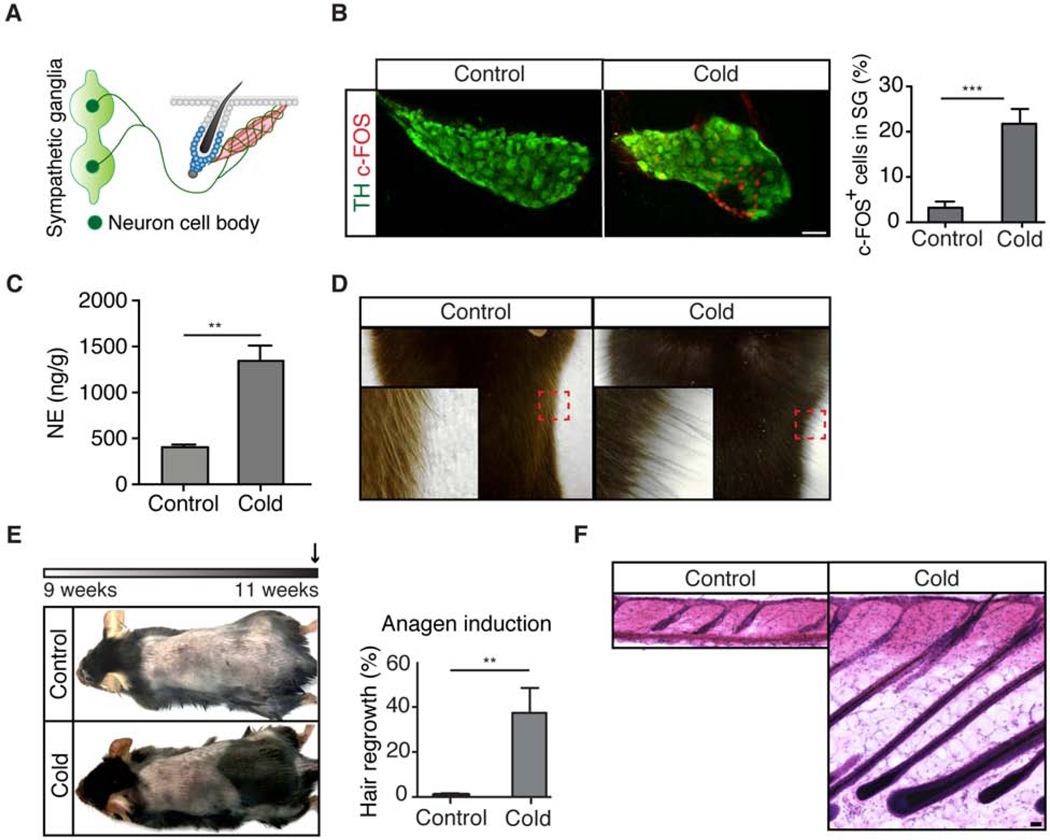
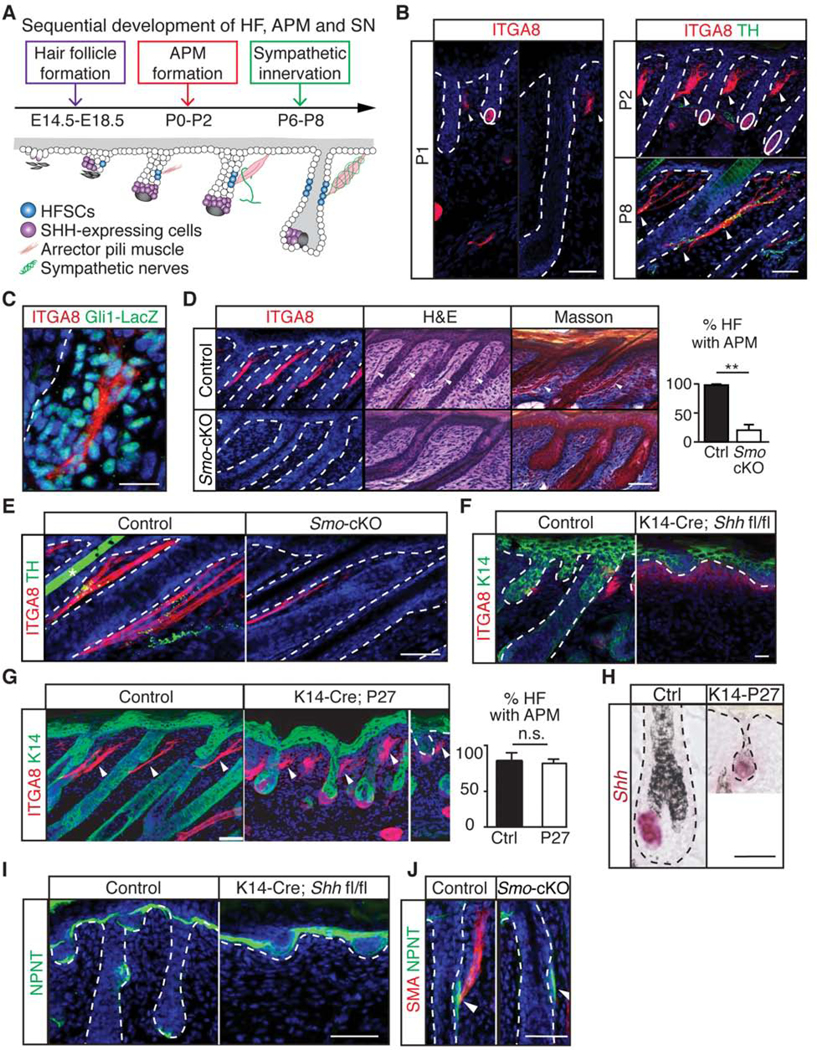
Piloerection (goosebumps) requires concerted actions of the hair follicle, the arrector pili muscle (APM), and the sympathetic nerve, providing a model to study interactions across epithelium, mesenchyme, and nerves. Here, we show that APMs and sympathetic nerves form a dual-component niche to modulate hair follicle stem cell (HFSC) activity. Sympathetic nerves form synapse-like structures with HFSCs and regulate HFSCs through norepinephrine, whereas APMs maintain sympathetic innervation to HFSCs. Without norepinephrine signaling, HFSCs enter deep quiescence by down-regulating the cell cycle and metabolism while up-regulating quiescence regulators Foxp1 and Fgf18. During development, HFSC progeny secretes Sonic Hedgehog (SHH) to direct the formation of this APM-sympathetic nerve niche, which in turn controls hair follicle regeneration in adults. Our results reveal a reciprocal interdependence between a regenerative tissue and its niche at different stages and demonstrate sympathetic nerves can modulate stem cells through synapse-like connections and neurotransmitters to couple tissue production with demands.
Keywords: Adrb2; hair follicle stem cells; nerve-stem-cell interaction; niche; stem cell quiescence; sympathetic nerve.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests A provisional patent application has been filed by the President and Fellows of Harvard College based on this work.
Figures
Comment in
-
A Hairy End to a Chilling Event.Cell. 2020 Aug 6;182(3):539-541. doi: 10.1016/j.cell.2020.07.004. Cell. 2020. PMID: 32763185
References
-
- Andl T, Reddy ST, Gaddapara T, and Millar SE (2002). WNT signals are required for the initiation of hair follicle development. Dev Cell 2, 643–653. - PubMed
-
- Asada-Kubota M. (1995). Inhibition of hair growth by subcutaneous injection of a sympathetic neurotoxin, 6-hydroxydopamine in neonatal mice. Anat Embryol (Berl) 191, 407–414. - PubMed
-
- Bai CB, Auerbach W, Lee JS, Stephen D, and Joyner AL (2002). Gli2, but not Gli1, is required for initial Shh signaling and ectopic activation of the Shh pathway. Development 129, 4753–4761. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
