

Translational Components Contribute to Acclimation Responses to High Light, Heat, and Cold in Arabidopsis
- PMID: 32679545
- PMCID: PMC7364123
- DOI: 10.1016/j.isci.2020.101331
Translational Components Contribute to Acclimation Responses to High Light, Heat, and Cold in Arabidopsis
Abstract
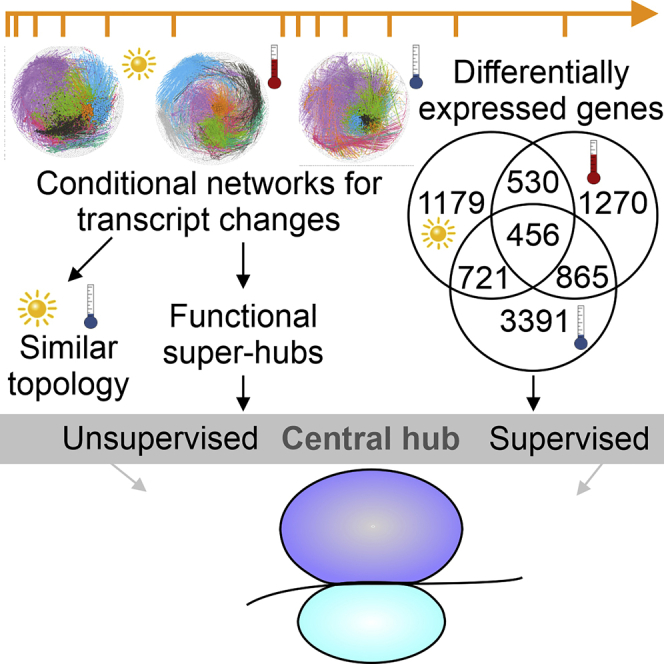
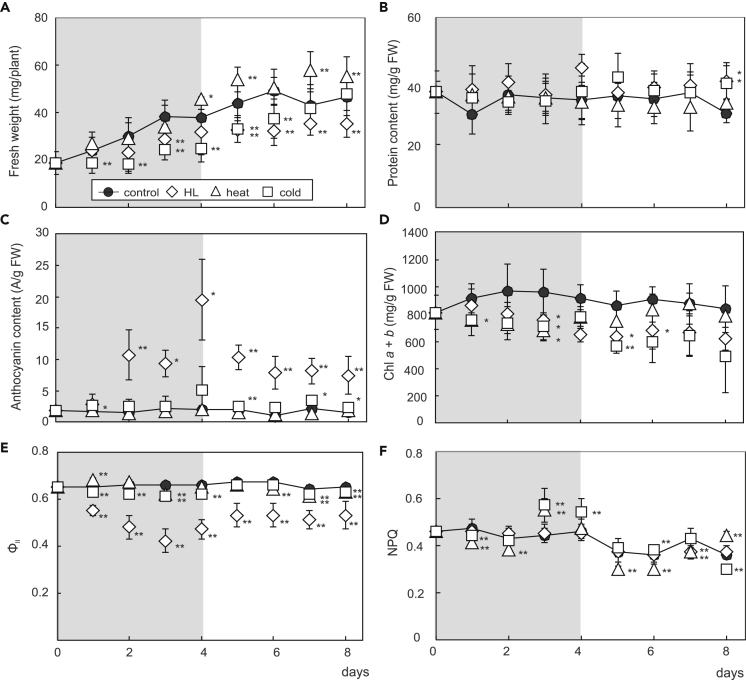
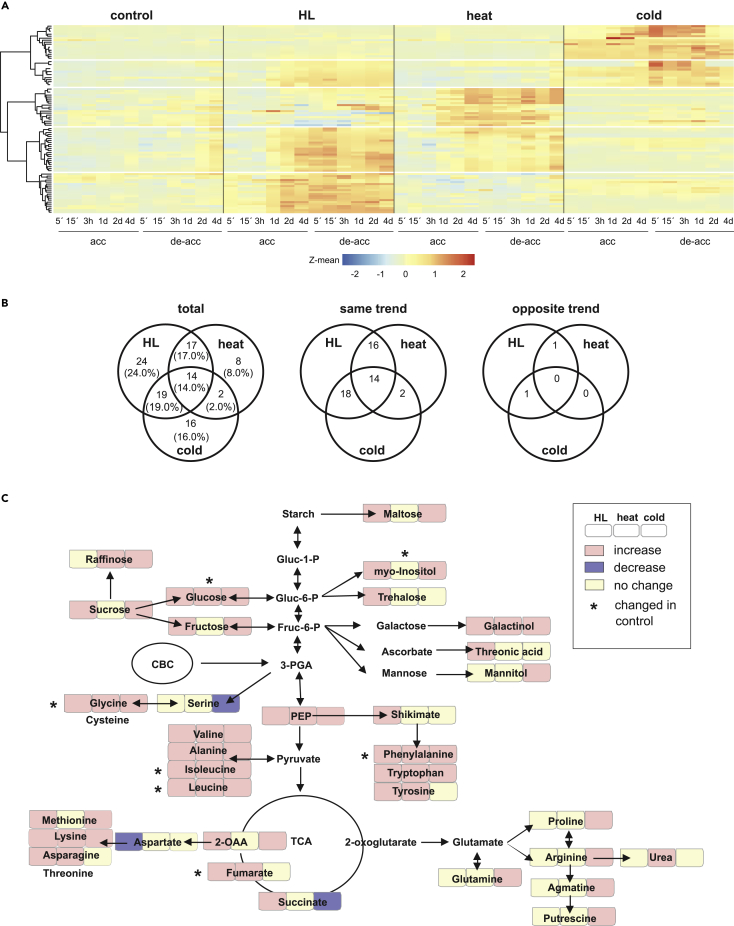
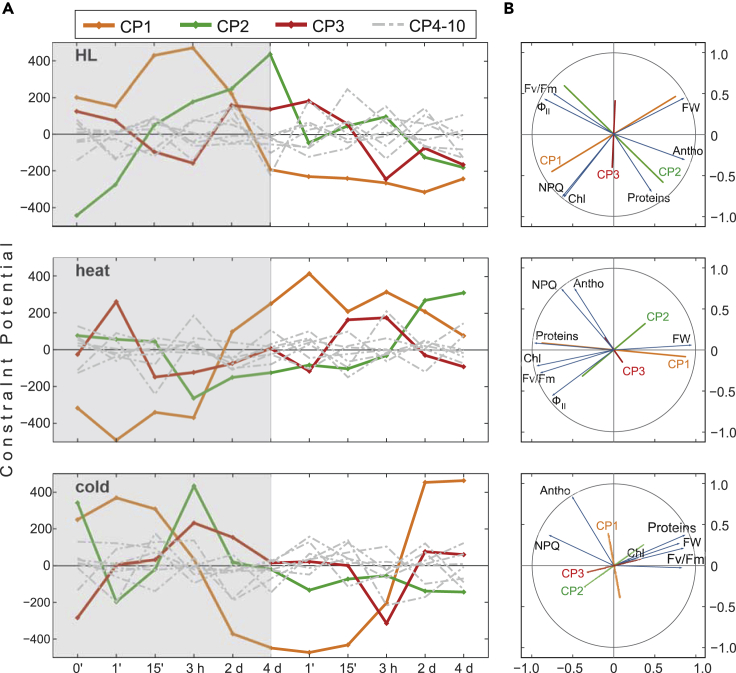
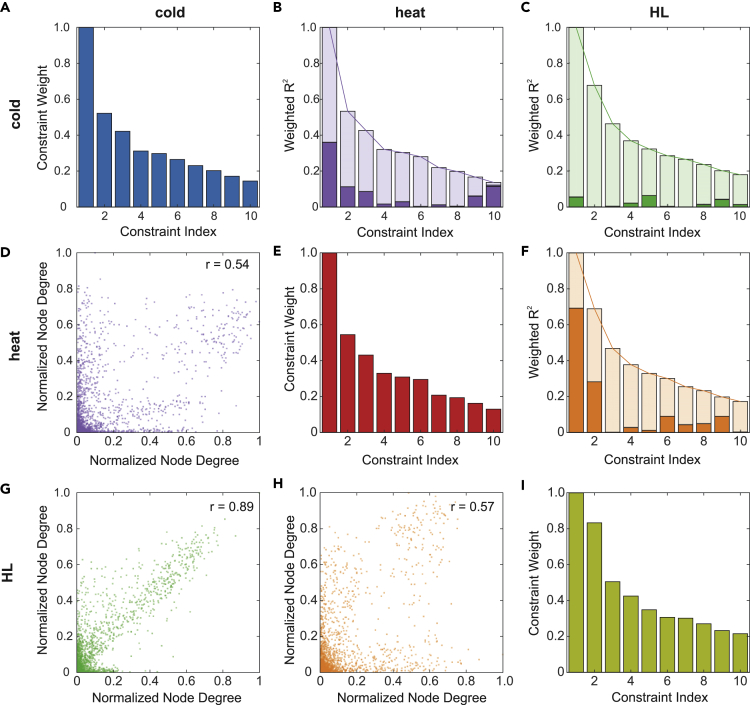
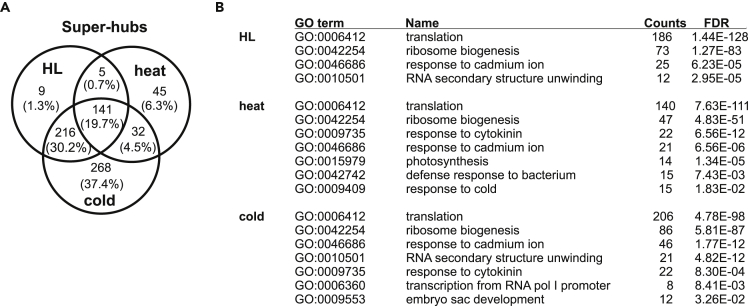
Plant metabolism is broadly reprogrammed during acclimation to abiotic changes. Most previous studies have focused on transitions from standard to single stressful conditions. Here, we systematically analyze acclimation processes to levels of light, heat, and cold stress that subtly alter physiological parameters and assess their reversibility during de-acclimation. Metabolome and transcriptome changes were monitored at 11 different time points. Unlike transcriptome changes, most alterations in metabolite levels did not readily return to baseline values, except in the case of cold acclimation. Similar regulatory networks operate during (de-)acclimation to high light and cold, whereas heat and high-light responses exhibit similar dynamics, as determined by surprisal and conditional network analyses. In all acclimation models tested here, super-hubs in conditional transcriptome networks are enriched for components involved in translation, particularly ribosomes. Hence, we suggest that the ribosome serves as a common central hub for the control of three different (de-)acclimation responses.
Keywords: Biological Sciences; Metabolomics; Plant Biology; Plants; Transcriptomics.
Copyright © 2020 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures

References
-
- Amme S., Matros A., Schlesier B., Mock H.P. Proteome analysis of cold stress response in Arabidopsis thaliana using DIGE-technology. J. Exp. Bot. 2006;57:1537–1546. - PubMed
-
- Anjum N.A. Plant acclimation to environmental stress: a critical appraisal. Front. Plant Sci. 2015;6:445.
-
- Ashraf M., Ahmad M.S.A., Öztürk M., Aksoy A. Crop improvement through different means: challenges and prospects. In: Ashraf M., Öztürk M., Ahmad M.S.A., Aksoy A., editors. Crop Production for Agricultural Improvement. Springer Netherlands; 2012. pp. 1–15.
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
