

Polymorphic Toxins and Their Immunity Proteins: Diversity, Evolution, and Mechanisms of Delivery
- PMID: 32680451
- PMCID: PMC8019152
- DOI: 10.1146/annurev-micro-020518-115638
Polymorphic Toxins and Their Immunity Proteins: Diversity, Evolution, and Mechanisms of Delivery
Abstract
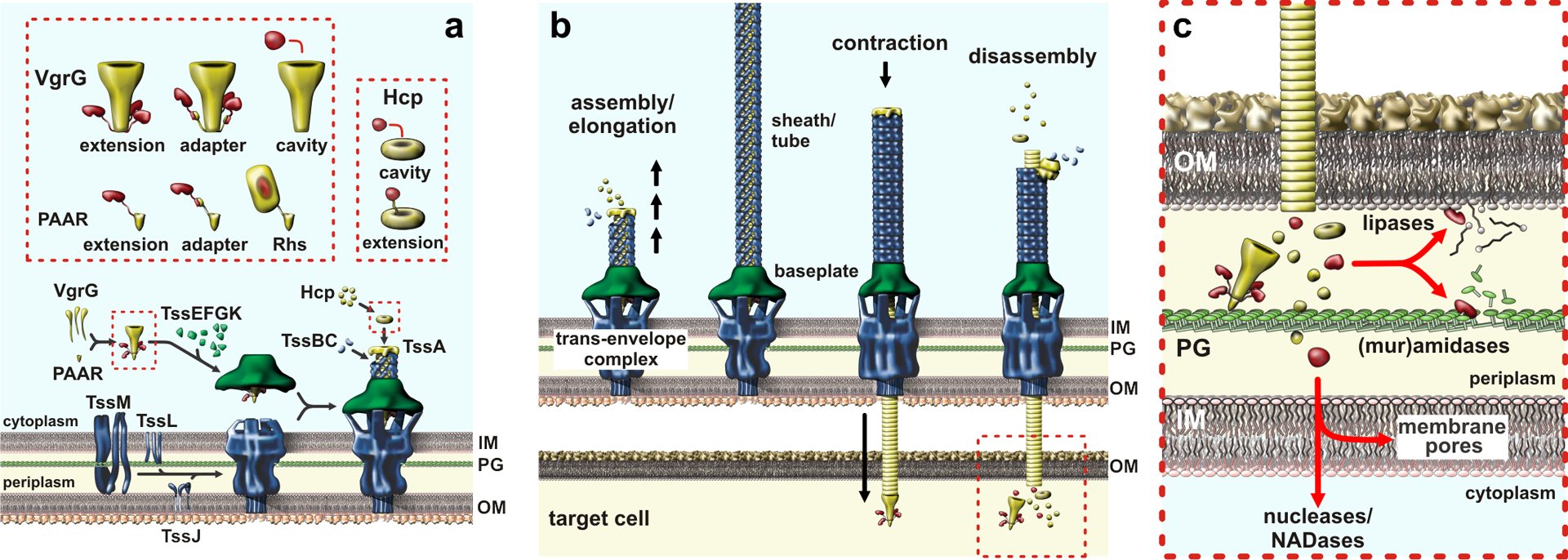
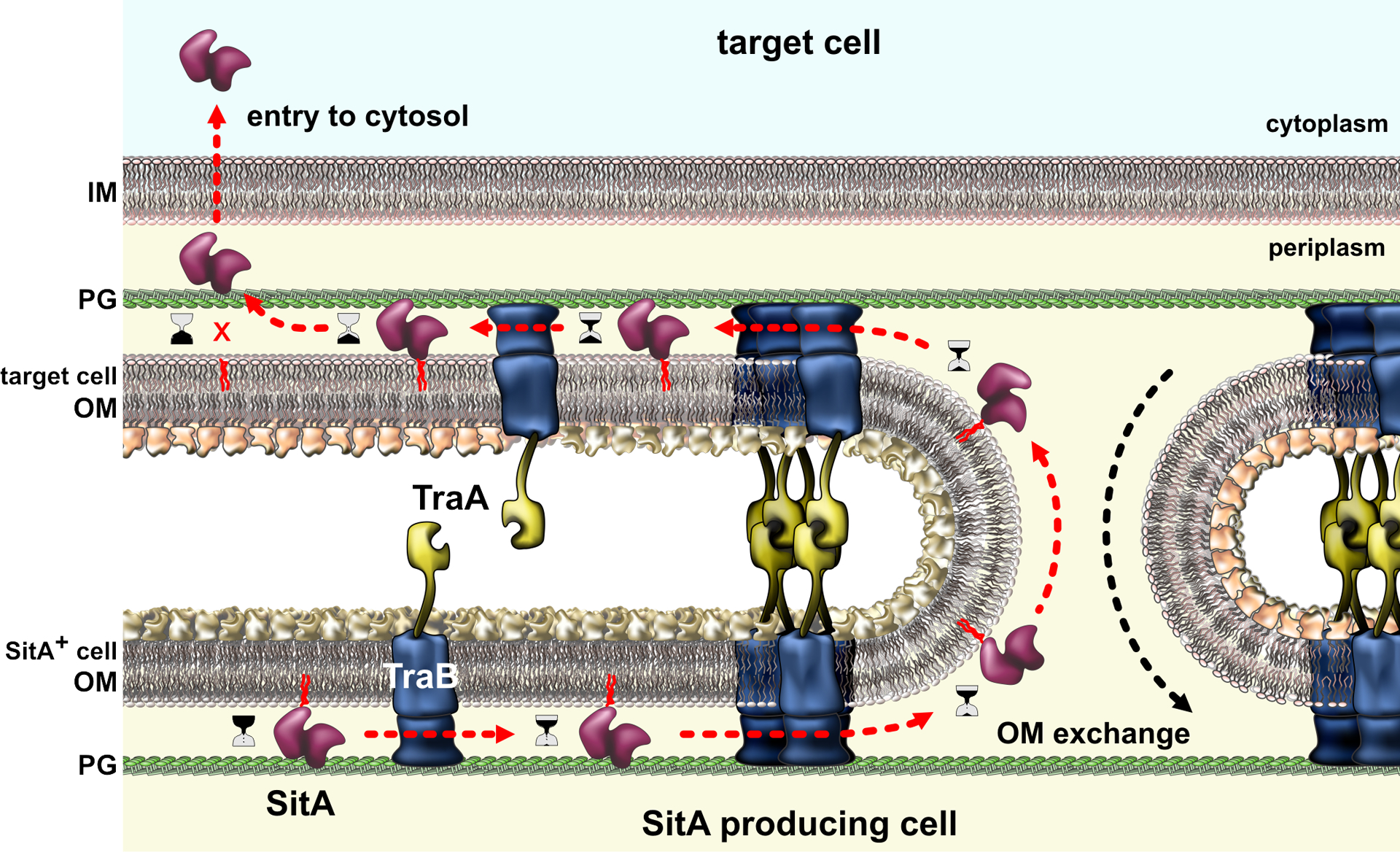
All bacteria must compete for growth niches and other limited environmental resources. These existential battles are waged at several levels, but one common strategy entails the transfer of growth-inhibitory protein toxins between competing cells. These antibacterial effectors are invariably encoded with immunity proteins that protect cells from intoxication by neighboring siblings. Several effector classes have been described, each designed to breach the cell envelope of target bacteria. Although effector architectures and export pathways tend to be clade specific, phylogenetically distant species often deploy closely related toxin domains. Thus, diverse competition systems are linked through a common reservoir of toxin-immunity pairs that is shared via horizontal gene transfer. These toxin-immunity protein pairs are extraordinarily diverse in sequence, and this polymorphism underpins an important mechanism of self/nonself discrimination in bacteria. This review focuses on the structures, functions, and delivery mechanisms of polymorphic toxin effectors that mediate bacterial competition.
Keywords: CDI; ESS; Esx-like secretion system; MafB; OME; T6SS; colicins; contact-dependent growth inhibition; outer membrane exchange; type VI secretion system.
Figures
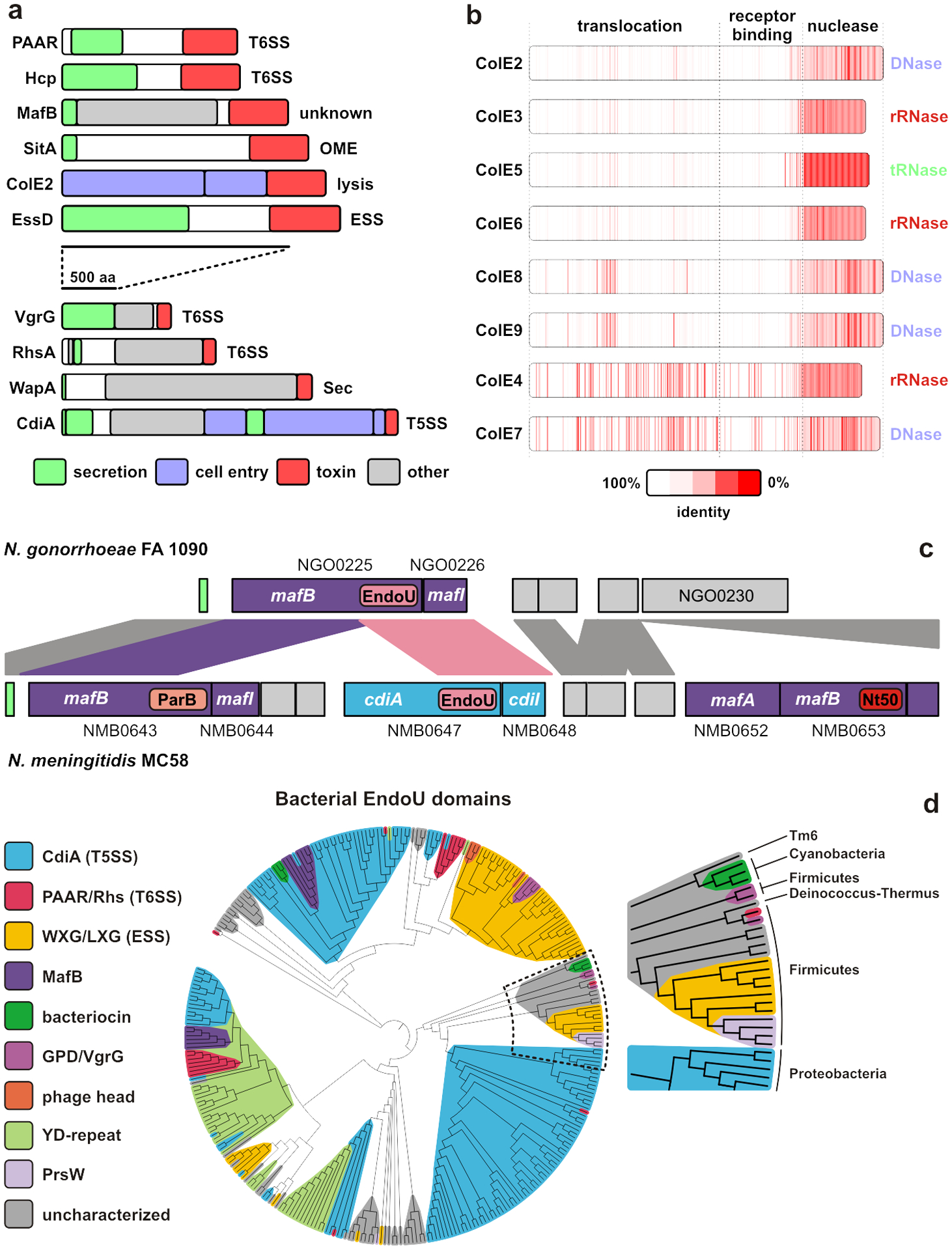
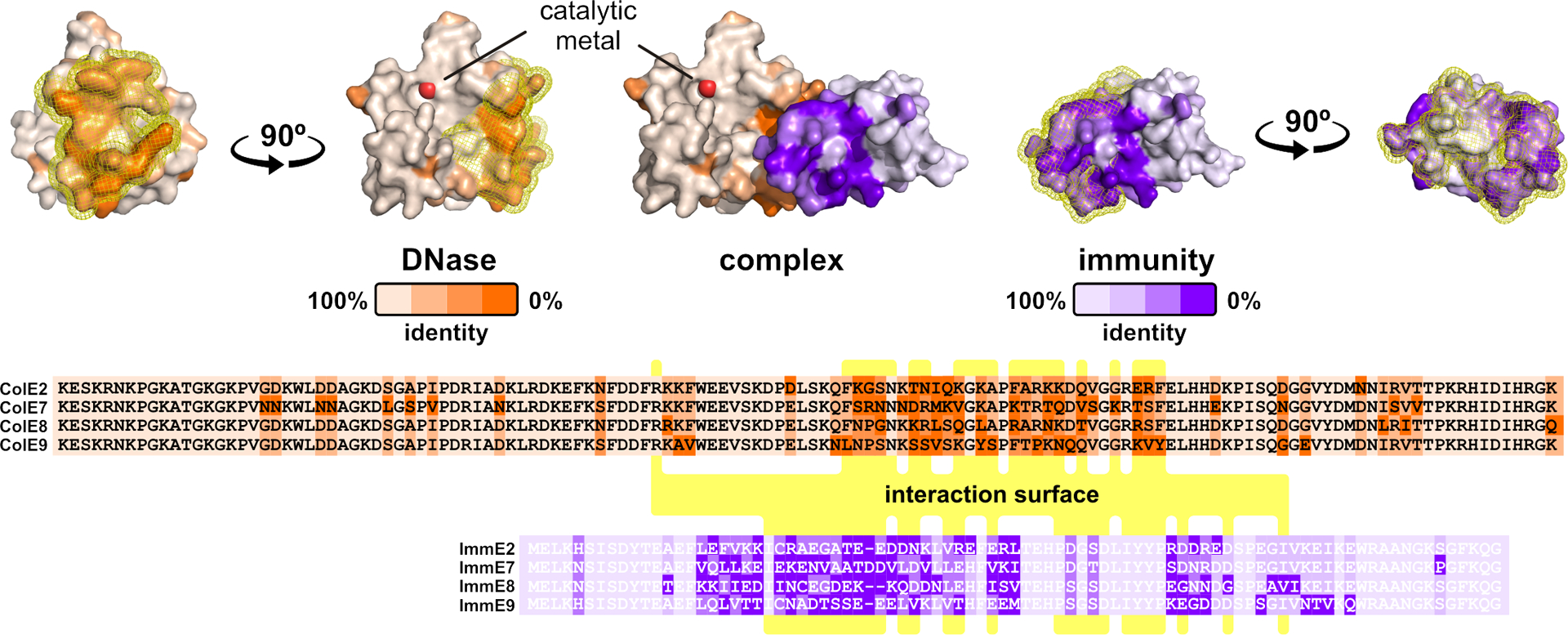
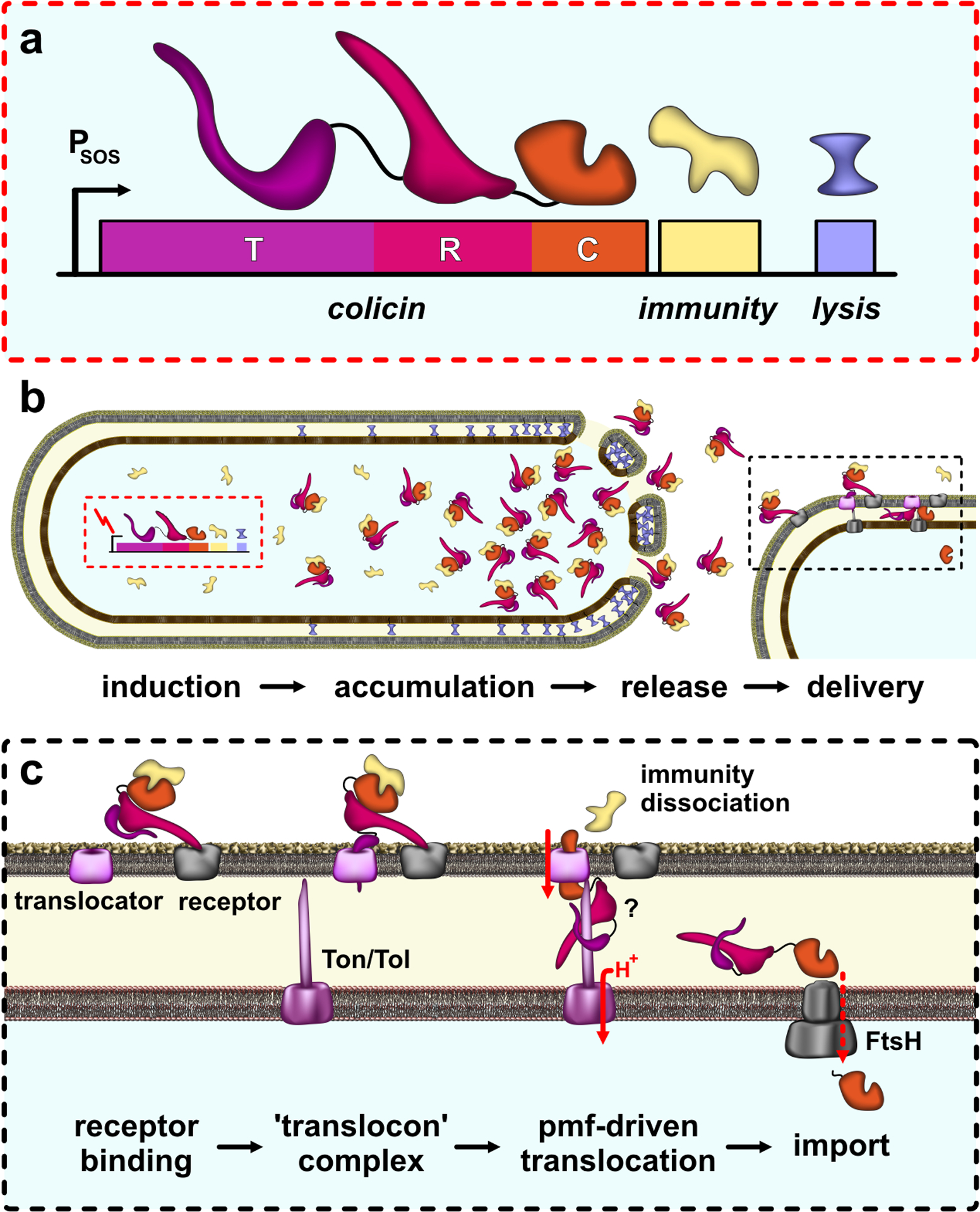
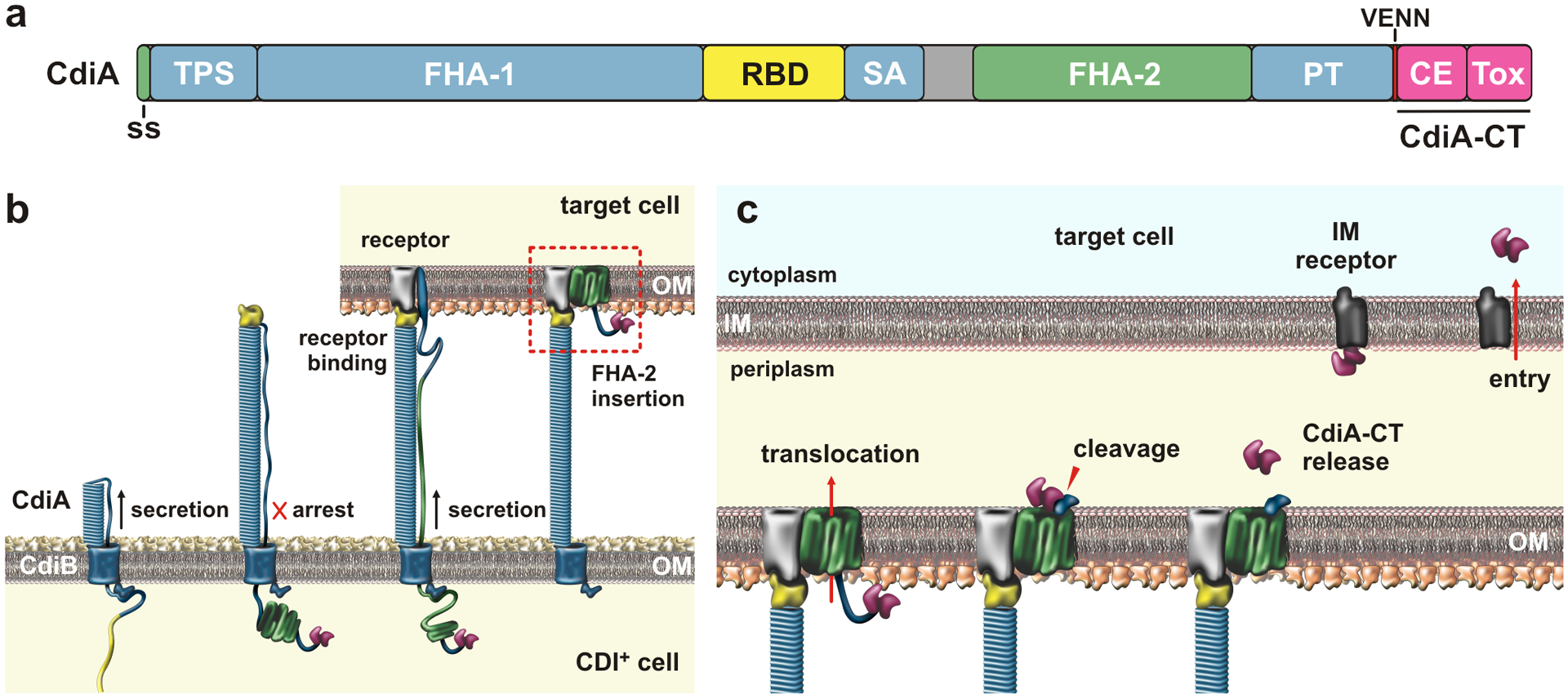
References
-
- Aoki SK, Pamma R, Hernday AD, Bickham JE, Braaten BA, Low DA. 2005. Contact-dependent inhibition of growth in Escherichia coli. Science 309:1245–8 - PubMed
