

Toxoplasma Mechanisms for Delivery of Proteins and Uptake of Nutrients Across the Host-Pathogen Interface
- PMID: 32680452
- PMCID: PMC9934516
- DOI: 10.1146/annurev-micro-011720-122318
Toxoplasma Mechanisms for Delivery of Proteins and Uptake of Nutrients Across the Host-Pathogen Interface
Abstract
Many intracellular pathogens, including the protozoan parasite Toxoplasma gondii, live inside a vacuole that resides in the host cytosol. Vacuolar residence provides these pathogens with a defined niche for replication and protection from detection by host cytosolic pattern recognition receptors. However, the limiting membrane of the vacuole, which constitutes the host-pathogen interface, is also a barrier for pathogen effectors to reach the host cytosol and for the acquisition of host-derived nutrients. This review provides an update on the specialized secretion and trafficking systems used by Toxoplasma to overcome the barrier of the parasitophorous vacuole membrane and thereby allow the delivery of proteins into the host cell and the acquisition of host-derived nutrients.
Keywords: Toxoplasma gondii; nutrient acquisition; parasitophorous vacuole; secreted effectors; trafficking; translocon.
Figures
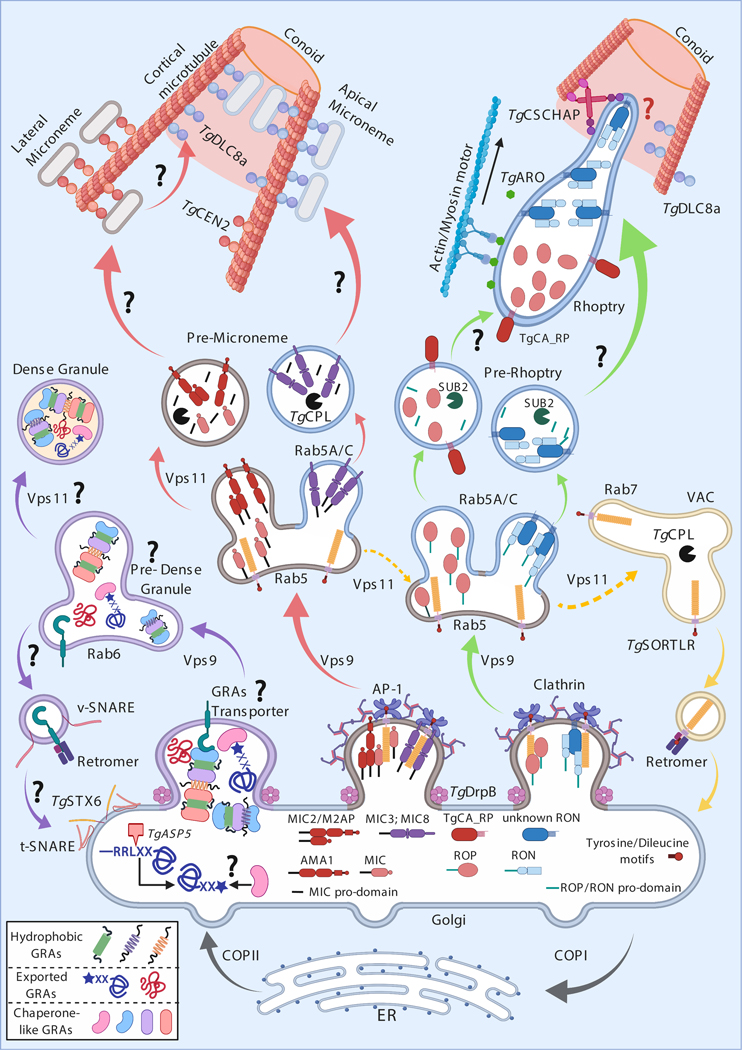
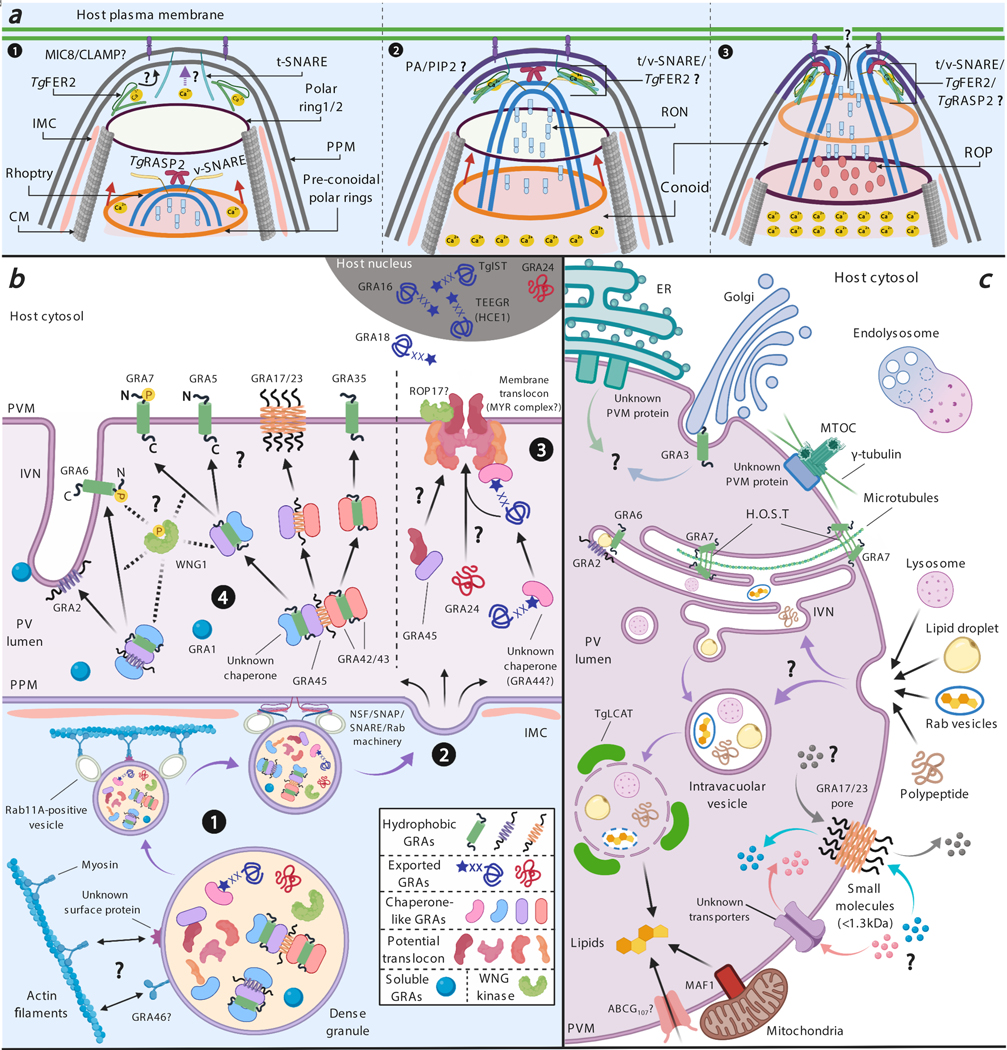
References
-
- Barlowe C, Orci L, Yeung T, Hosobuchi M, Hamamoto S, et al. 1994. COPII: a membrane coat formed by Sec proteins that drive vesicle budding from the endoplasmic reticulum. Cell. 77(6):895–907 - PubMed
-
- Bermudes D, Peck KR, Afifi MA, Beckers CJ, Joiner KA. 1994. Tandemly repeated genes encode nucleoside triphosphate hydrolase isoforms secreted into the parasitophorous vacuole of Toxoplasma gondii. J. Biol. Chem 269(46):29252–60 - PubMed
