

Molecular characterization and gene expression modulation of the alternative oxidase in a scuticociliate parasite by hypoxia and mitochondrial respiration inhibitors
- PMID: 32681023
- PMCID: PMC7367826
- DOI: 10.1038/s41598-020-68791-9
Molecular characterization and gene expression modulation of the alternative oxidase in a scuticociliate parasite by hypoxia and mitochondrial respiration inhibitors
Abstract
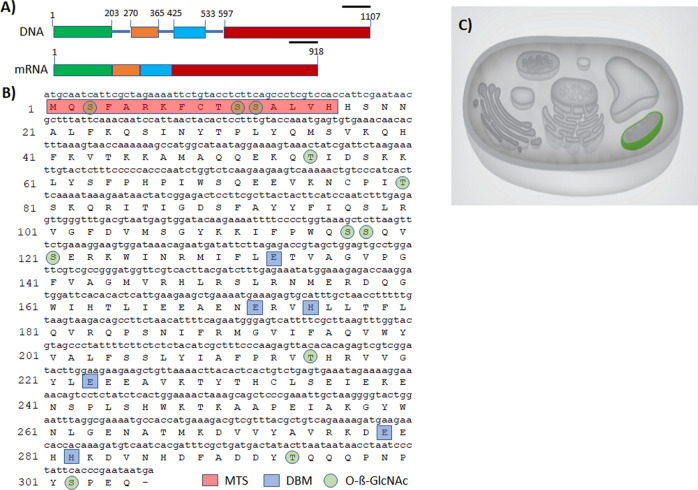
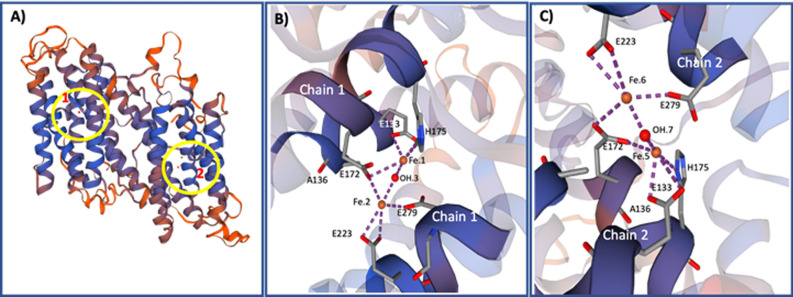
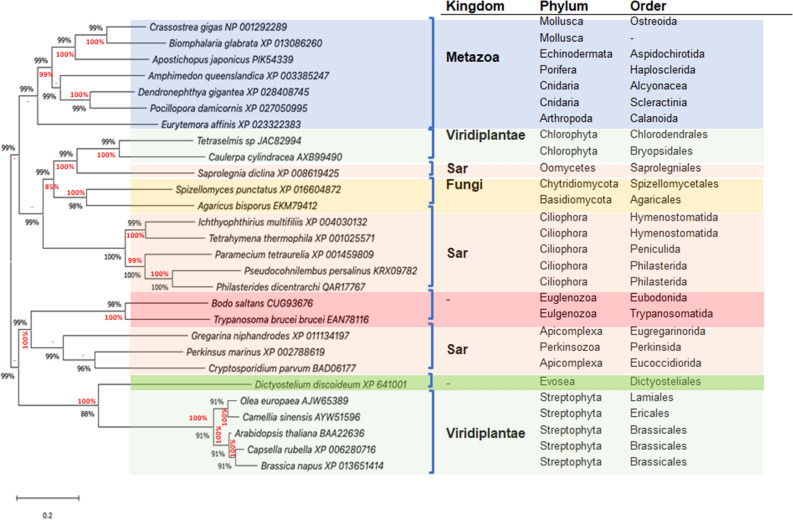
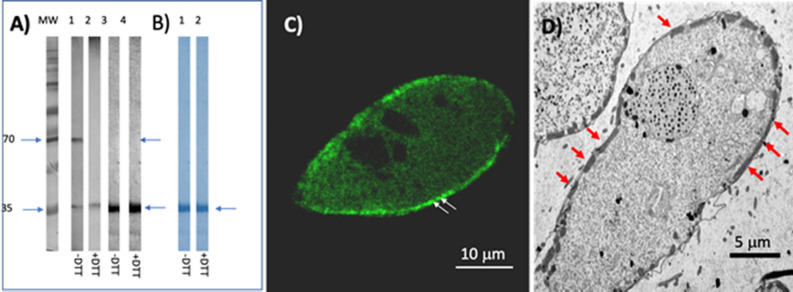
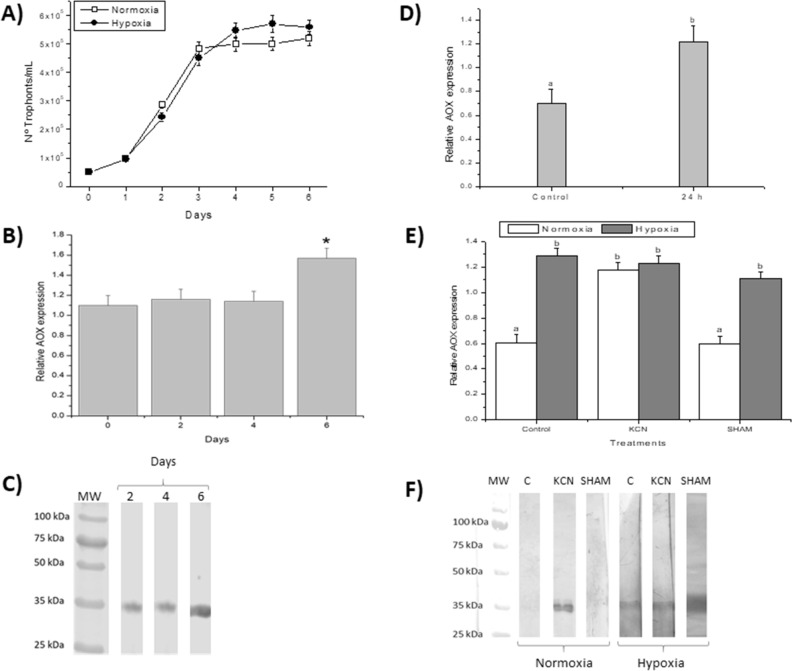
Philasterides dicentrarchi is a marine benthic microaerophilic scuticociliate and an opportunistic endoparasite that can infect and cause high mortalities in cultured turbot (Scophthalmus maximus). In addition to a cytochrome pathway (CP), the ciliate can use a cyanide-insensitive respiratory pathway, which indicates the existence of an alternative oxidase (AOX) in the mitochondrion. Although AOX activity has been described in P. dicentrarchi, based on functional assay results, genetic evidence of the presence of AOX in the ciliate has not previously been reported. In this study, we conducted genomic and transcriptomic analysis of the ciliate and identified the AOX gene and its corresponding mRNA. The AOX gene (size 1,106 bp) contains four exons and three introns that generate an open reading frame of 915 bp and a protein with a predicted molecular weight of 35.6 kDa. The amino acid (aa) sequence of the AOX includes an import signal peptide targeting the mitochondria and the protein is associated with the inner membrane of the mitochondria. Bioinformatic analysis predicted that the peptide is a homodimeric glycoprotein, although monomeric forms may also appear under native conditions, with EXXH motifs associated with the diiron active centers. The aa sequences of the AOX of different P. dicentrarchi isolates are highly conserved and phylogenetically closely related to AOXs of other ciliate species, especially scuticociliates. AOX expression increased significantly during infection in the host and after the addition of CP inhibitors. This confirms the important physiological roles of AOX in respiration under conditions of low levels of O2 and in protecting against oxidative stress generated during infection in the host.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Moore AL, Albury MS. Further insights into the structure of the alternative oxidase: From plants to parasites. Biochem. Soc. Trans. 2008;36:1022–1026. - PubMed
-
- Albury MS, Elliott C, Moore AL. Towards a structural elucidation of the alternative oxidase in plants. Physiol. Plant. 2009;137:316–327. - PubMed
-
- Ünlü ES, Ünüvar ÖC, Aydın M. Identification of alternative oxidase encoding genes in Caulerpa cylindracea by de novo RNA-Seq assembly analysis. Mar. Genom. 2019;46:41–48. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
